
{kind=link}
Tyrannosaurinae
|
|
Cet article ou cette section d'un article de zoologie doit être recyclé. Une réorganisation et une clarification du contenu est nécessaire.
Discutez des points à améliorer en page de discussion.
|
|
|
La mise en forme de cet article est à améliorer. (novembre 2014). La mise en forme du texte ne suit pas les recommandations de Wikipédia. La typographie, les liens internes ou externes, les conventions de style, la présentation des sources, etc. sont autant de points qui ne peuvent pas convenir voire être absents.
Ces motifs sont peut-être précisés sur la page de discussion. — Découvrez comment faire pour le « wikifier ». |

Silhouette d'un Tyrannosauriné.
† Tyrannosaurinae
— auteur incomplet —, date à préciser
Genres de rang inférieur
- Tyrannosaurus rex
- Alioramus ?
- Aublysodon (nomen dubium)
- Bistahieversor ?
- Daspletosaurus
- Deinodon (nomen dubium)
- Lythronax
- Nanotyrannus (nomen dubium)
- Nanuqsaurus
- Tarbosaurus
- Teratophoneus
- Tyrannosaurus
- Zhuchengtyrannus
Les Tyrannosaurinae sont une sous-famille de dinosaures de la famille des Tyrannosaures.
Liste des genres
- Alioramus
- Aublysodon ?
- Bistahieversor
- Daspletosaurus
- Deinodon ?
- Lythronax
- Nanotyrannus ?
- Nanuqsaurus
- Tarbosaurus
- Teratophoneus
- Tyrannosaurus
- Zhuchengtyrannus
Classification
| Tyrannosauridae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
Paléobiologie

Paléopathologie
Des scientifiques ont découvert dans les fossiles de T. Rex, des traces de goutte. Ce grand carnivore devait donc souffrir de douleurs aiguës et soudaines dans les articulations, ce qui devait probablement jouer sur son comportement[1].
Longévité
L'identification de plusieurs spécimens de jeunes Tyrannosaurus rex a permis aux scientifiques de documenter les changements ontogénétiques, d'évaluer le cours de leur vie, et de déterminer la croissance de ces animaux. Le plus petit individu connu (LACM 28471, le théropode « Jordan ») devait peser seulement 29,9 kg, alors que les plus grands, comme le spécimen FMNH PR2081 (Sue), devaient bien atteindre plus de 5 400 kg (soit presque 6 tonnes). Des analyses histologiques d'os de T. rex ont démontré que le spécimen LACM 28471 était âgé de deux ans seulement, alors que Sue avait 28 ans, probablement l'âge limite de cette espèce[2].
L'histologie a également permis de déterminer l'âge d'autres spécimens. Il est possible de dessiner les courbes de croissance lorsqu'on peut reporter sur un graphique l'âge et le poids de différents spécimens. Celle du T. rex est une courbe en S, les jeunes ne dépassaient pas les 1 800 kg jusqu'à 14 ans environ, puis leur taille augmentait de façon significative. Durant cette phase de croissance rapide, un jeune T. rex pouvait gagner en moyenne 600 kg par an pendant quatre ans. À partir de 18 ans, la courbe se stabilise, ce qui signifie que la croissance de l'animal augmente plus lentement. Par exemple, seulement 600 kg séparent le spécimen Sue âgé de 28 ans du spécimen canadien âgé quant à lui de 22 ans (RTMP 81.12.1)[2]. Une autre étude histologique plus récente menée par différents scientifiques corrobore ces résultats, démontrant que la croissance rapide commence à ralentir à partir de 16 ans[3]. Cette cassure brutale de la vitesse de croissance entre 14 et 18 ans pourrait témoigner de l'existence d'une phase de maturité physique, synonyme de maturité sexuelle. Une hypothèse soutenue par la découverte en 2005 de tissus osseux riches en calcium, connus aussi sous le nom d'os médullaires, dans le fémur d'une T. rex âgée de 18 ans (MOR 1125, également connu sous le nom de « B-rex »)[4]. Dans la nature, ces tissus osseux ne sont retrouvés que chez les oiseaux femelles matures juste avant la ponte; ils permettent de renforcer la coquille des œufs. Les dinosaures étant également ovipares, le même phénomène intervenait peut-être à l'époque[5],[6] et cette découverte pourrait donc indiquer que la jeune « B-rex » était sexuellement mature[7]. La même découverte a été réalisée chez une femelle Allosaurus âgée de 10 ans et chez une femelle Tenontosaurus âgée de 8 ans, laissant penser que la maturité sexuelle des dinosaures serait intervenue beaucoup plus tôt que ce qui était jusqu'alors pensé. Des femelles dinosaures pouvaient donc pondre des œufs dès leur préadolescence, et de là, devenir mères[6],[8]. D'autres tyrannosauridés ont des courbes de croissances très similaires à celles de T. rex, même si leurs vitesses de croissance plus lentes correspondent à des tailles plus petites à l'âge adulte[9].
Le taux de mortalité augmente à l'approche de la maturité sexuelle, un schéma que l'on retrouve chez les autres tyrannosaures, les grands oiseaux et les mammifères. Ces espèces connaissent après une mortalité infantile élevée, un taux de survivance élevé, qui décline rapidement à la maturité sexuelle. La mortalité augmente à la maturité sexuelle, en partie dû au stress des femelles qui accompagne la ponte. Une étude démontre que le faible nombre d'échantillons de petits est dû en partie à leur taux de mortalité bas, ces animaux ne mouraient pas en grand nombre à ces âges[9].
Vision

Lythronax avait un museau court et étroit, un large crâne à l'arrière et les yeux orientés vers l'avant. Grâce à cette anatomie, le dinosaure devait avoir une vision stéréoscopique et percevoir ainsi la profondeur. T. rex avait une anatomie semblable selon Loewen[10]. De plus, l'arrière ramassée de sa tête lui permettait d'avoir une vision étendue[11].
Dimorphisme sexuel

Avec la découverte de spécimens de plus en plus nombreux, les scientifiques ont commencé à analyser les différences entre individus et ils ont identifié chez Tyrannosaurus rex, deux types morphologiques distincts, similaires à d'autres espèces de théropodes. En prenant en compte leur morphologie générale, l'un a été dénommé le morphotype « robuste » alors que l'autre était dénommé « gracile ».
Plusieurs caractéristiques associées à ces deux morphotypes ont permis aux scientifiques d'en déduire qu'il s'agissait peut-être d'un dimorphisme sexuel, et que le type robuste était probablement femelle, alors que le type gracile était probablement mâle. Ainsi le pelvis de plusieurs spécimens robustes est plus large, peut-être afin de faciliter le passage des œufs dans le bassin lors de la ponte[12]. De plus chez le type robuste, on retrouve un arc hémal ou chevron — os protégeant certains éléments vitaux de la partie ventrale de la queue — de taille réduite au niveau de la première vertébrale caudale, peut-être également afin de faciliter le passage des œufs dans le conduit génital[13]. Cette hypothèse avait été déjà proposée par Romer en 1956 pour les crocodiliens[14].
Au début des années 2000, l'existence d'un dimorphisme sexuel chez T. rex a été remise en question. Ainsi en 2005, une étude concluait que la présence de chevrons ne permettait pas de différencier le genre des crocodiliens, et jetait ainsi le doute sur le bien-fondé de ce critère pour différencier le genre de T. rex[15]. De plus les scientifiques ont découvert que le premier chevron de Sue, un spécimen extrêmement robuste, était en fait situé très près de son pelvis, comme chez certains reptiles mâles, démontrant que la position des chevrons variait trop pour être un bon indicateur du sexe, chez les T. rex comme chez les reptiles modernes[16]. L'existence de différences morphologiques entre les différents spécimens retrouvés pourrait ne pas être liée à un dimorphisme sexuel, mais plutôt à des variations géographiques, ou à des variations dues à l'âge, les individus robustes étant les plus âgés[17].

En 2009, nous ne connaissons avec certitude le sexe que d'un seul et unique spécimen de T. rex. L'examen de « B.rex » (MOR 1125) a permis de retrouver des tissus mous préservés provenant d'os médullaire, un tissu spécialisé retrouvé chez les oiseaux femelles modernes, source de calcium permettant la production de la coquille des œufs lors de l'ovulation. Le tissu osseux médullaire n'est retrouvé naturellement que chez les femelles oiseaux, suggérant fortement que « B.rex » était une femelle, et qu'elle est morte en période ovulatoire[7]. Des études récentes ont montré que le tissu médullaire n'était jamais retrouvé chez les crocodiles, qui sont les animaux vivants les plus proches des dinosaures, avec les oiseaux. La présence partagée de tissu médullaire chez les oiseaux et les dinosaures théropodes montre les liens évolutionnaires étroits entre les deux[18].
Alioramus avait une tête plus longue et plus petite et des dents plus nombreuses que les autres tyrannosaures. Il était petit et svelte et possédait six petites cornes sur son long museau, une de chaque côté et quatre au milieu en une seule rangée. Trop petites pour se défendre, ces cornes pourraient avoir été des caractéristiques sexuelles secondaires, identifiant les mâles et les femelles.
Croissance
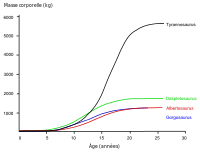
Les paléontologues Gregory Erickson et des collègues ont étudié la croissance et le développement des tyrannosauridés. Des analyses de l'histologie des os peuvent déterminer l'âge d'un spécimen à sa mort. Les taux de croissance peuvent être examinés quand les âges de différents individus sont comparés à leurs masses sur un graphe. Erickson a montré qu'après une longue période en tant que juvéniles, les tyrannosaures subissent des poussées de croissance importantes pendant environ quatre ans, vers la moitié de leur vie. La phase de croissance rapide se termine par la maturité sexuelle. Erickson a seulement examiné les Daspletosaurus de la formation de Dinosaur Park, mais ces spécimens montrent le même modèle. Comparé aux albertosaurinés, les tyrannosaurinés montrent un taux de croissance plus rapide, dû à leur taille adulte plus importante. Le taux de croissance le plus important pour les Daspletosaurus est 180 kilogrammes par an, selon une estimation de la masse de 1800 kilogrammes pour un adulte. D'autres auteurs ont suggéré des poids plus importants pour les Daspletosaurus adultes; ce qui changerait la magnitude du taux de croissance mais pas le modèle dans son ensemble[2].
Par la tabulation du nombre de spécimens de chaque groupe d'âge, Erickson et ses collègues purent tirer des conclusions sur l'histoire d'une population d'Albertosaurus. Leur analyses ont montré que bien que les juvéniles soient rares dans l'enregistrement fossile, les individus subadultes en phase de croissance rapide et les adultes sont beaucoup plus courants. Cela pourrait être des biais dus à la préservation ou à la récolte, mais Erickson émit l'hypothèse que la différence était due à une faible mortalité des individus juvéniles au-delà d'une certaine taille, ce qui s'observe chez des grands mammifères actuels comme l'éléphant. Cette faible mortalité peut résulter d'un manque de prédation, puisque les tyrannosaures surpassaient tous les prédateurs contemporains en taille dès leur deuxième année. Les paléontologues n'ont pas trouvé assez de restes de Daspletosaurus pour faire une analyse similaire, mais Erickson note que la même tendance générale semble s'appliquer[9].
Comportement
Chasseurs ou charognards
Jack Horner s'est demandé si les tyrannosaures étaenit des chasseurs ou des charognards. En effet, de ne sont pas de bons coureurs. Cependant, ils auraient eu un flair très sensible, compensant ainsi leur mauvaise vision : cela leur aurait donc permis de repérer facilement une charogne. Néanmoins, on a découvert une morsure de tyrannosaure sur un edmontosaurus qui avait cicatrisé ; l'animal est décédé postérieurement. De plus le tyrannosaure détenait une mâchoire d'une puissance phénoménale, qu'il devait probablement utiliser pour tuer des animaux vivants, ce qui en ferait un chasseur.
Les scientifiques se demandent donc toujours s'il mangeait de la viande fraîche ou de la charogne. Il n'est d'ailleurs pas exclu qu'il ait pu être à la fois prédateur et charognard, selon les opportunités qui se présentaient à lui. À titre de comparaison, dans les comportements que l'on peut observer de nos jours, il peut arriver que les vautours par exemple, des charognards, chassent lorsque la faim les y pousse. À l'inverse, les lions, découvrant un cadavre encore frais, n'hésitent pas à s'en repaître. Ils s'avèrent par là, opportunistes.
Vie en groupe
Le paléontologue Jack Horner a découvert cinq tyrannosaures dans un même gisement. Il suppose qu'ils vivaient en groupe. Ces individus n'ont pas pu être attirés par une proie ou une charogne : aucun reste d'herbivore n'ayant été trouvé à cet endroit.
Un jeune spécimen des espèces de Daspletosaurus de la formation de Dinosaur Park (TMP 94.143.1) présente des traces de morsure sur le crâne, infligées par un autre tyrannosaure. Les marques ont guéri, ce qui indique que l'animal a survécu à la morsure. Un Daspletosaurus adulte de cette même formation (TMP 85.62.1) montre des marques similaires de morsure d'un tyrannosaure, ce qui montre que ces attaques n'étaient pas dirigées uniquement contre les jeunes. Bien que ces morsures puissent être attribuées à d'autres espèces, l’agression intraspécifique, dont la morsure faciale, est très commune parmi les prédateurs. Ces morsures à la face se retrouvent chez d'autres tyrannosaures comme Gorgosaurus et Tyrannosaurus, ainsi que dans d'autres groupes de théropodes comme Sinraptor et Saurornitholestes. Darren Tanke et Phil Currie émirent l'hypothèse que les morsures sont dues à la compétition intraspécifique pour le territoire ou les ressources, ou pour la dominance dans un groupe social[19].
Des preuves que Daspletosaurus vivait en groupes sociaux ont été trouvées dans un niveau riche en os de la formation Two Medicine dans le Montana. Il contenait les restes de trois Daspletosaurus, dont un adulte, un juvénile, et un autre individu d'une taille intermédiaire. Au moins cinq hadrosaures sont préservés au même endroit. La géologie de la région prouve que les restes n'ont pas été charriés pas un cours d'eau mais que tous ces animaux ont été ensevelis au même endroit simultanément. Les restes des hadrosaures sont dispersés et portent de nombreuses marques de dents de tyrannosaure, ce qui indique que les Daspletosaurus étaient en train de se nourrir des hadrosaures au moment de la mort. La cause de la mort est inconnue. Currie imagine que les dasplétosaures formaient une meute, bien que cela ne puisse pas être affirmé de façon certaine[20]. D'autres scientifiques restent sceptiques quant aux preuves des groupes sociaux de Daspletosaurus et d'autres grands théropodes[21]. Brian Roach et Daniel Brinkman ont suggéré que les interactions sociales de Daspletosaurus devaient ressembler à celles du dragon de Komodo, où des individus non-coopératifs se massent sur des carcasses, s'attaquant souvent parfois même allant jusqu'au cannibalisme[22].
Paléoécologie
Coexistance entre Daspletosaurus et Gorgosaurus
À la fin du Campanien, en Amérique du Nord, Daspletosaurus était contemporain du tyrannosauridé de la sous-famille des Albertosaurinae Gorgosaurus. C'est une des rares coexistences de genres de tyrannosaures connues. Dans les guildes de prédateurs modernes, des prédateurs de tailles similaires sont séparés dans des niches écologiques différentes par des caractères anatomiques, comportementaux ou géographiques différents qui limitent la compétition[23]. Plusieurs études ont tenté d'expliquer la différenciation des niches écologiques de Daspletosaurus et Gorgosaurus.
Dale Russell émit l'hypothèse que Gorgosaurus, moins solide et plus commun, devait chasser les hadrosaures, tandis que Daspletosaurus, plus robuste et moins commun, devait s'être spécialisé dans la chasse des cératopsidés, moins abondants mais avec une meilleure défense, qui les aurait rendus plus difficiles à chasser[24]. Cependant, un spécimen de Daspletosaurus (OTM 200) de la formation Two Medicine contient les restes d'un hadrosaure juvénile dans sa cavité abdominale[25]. Les mâchoires plus grandes et plus larges des tyrannosaurinés comme Daspletosaurus sont plus fortes que celles des albertosaurinés comme Gorgosaurus, bien que la force des dents des deux groupes soit similaire. Cela semble montrer une différence dans le mode d'alimentation ou le régime alimentaire[26]. D'autres auteurs ont suggéré que la compétition était limitée par une séparation géographique. À la différence d'autres groupes de dinosaures, il semble qu'il n'y ait pas de corrélation avec la distance de la mer. Ni Daspletosaurus ni Gorgosaurus était plus abondant que l'autre à des altitudes hautes ou basses[23]. Cependant, bien qu'il y ait un chevauchement, Gorgosaurus semble plus commun vers le nord, et les espèces de Daspletosaurus plus abondantes dans le sud. Ce modèle se retrouve dans d'autres groupes de dinosaures. Les cératopsiens cératopsinés et les hadrosaures hadrosaurinés sont aussi plus communs dans la formation Two Medicine et dans le sud-ouest de l'Amérique du Nord pendant le Campanien. Thomas Holtz pense que ces modèles peuvent montrer des préférences écologiques partagées entre les tyrannosaurinés, cératopsinés et hadrosaurinés. Holtz note qu'à la fin du Maastrichtien, les tyrannosaurinés comme Tyrannosaurus rex, les hadrosaurinés et les cératopsinés comme Triceratops étaient répandus à travers l'ouest de l'Amérique du Nord, alors que les albertosaurinés et les centrosaurinés étaient éteints, et les lambeosaurinés étaient très rares[27].
Synonymie entre Tyrannosaurus et Tarbosaurus ?
Le Tarbosaurus possédait une anatomie proche de celle de son cousin tyrannosaure, ce fut la cause de cette question qui sera débattue par nombre de savants; « Le tarbosaurus et le tyrannosaure ne feraient-ils qu'un ? ». En effet les deux reptiles possédaient la même anatomie, tous deux ayant les membres antérieurs atrophiés (pour équilibrer leur poids de part et d'autre des appuis au sol, car ils possédaient une tête volumineuse), et tous deux ayant la même dentition, la même ossature et une taille sensiblement semblable (10 à 14 mètres contre 12 à 15 mètres pour le Tyrannosaurus rex).
Cependant, les yeux du tyrannosaure sont disposés de façon plus frontale que ceux du tarbosaurus qui a un crâne plus étroit[28]. On considère aujourd'hui Tarbosaurus comme étant plus proche d'Alioramus que de Tyrannosaurus[réf. nécessaire].
Classification et systématique
Carr et al. 2005[29]
*Note: Carr et al. utilisent le nom Tyrannosaurus bataar Currie et al. 2003[30]
Cladogrammes des Tyrannosaurinae |
Daspletosaurus est généralement considéré comme étant proche parent de Tyrannosaurus rex, ou même un ancêtre direct par anagénèse[31]. Gregory Paul réassigna D. torosus au genre Tyrannosaurus, créant ainsi le Tyrannosaurus torosus[32], mais cela n'a pas été encore généralement accepté[33],[27]. Beaucoup de chercheurs pensent que les genres Tarbosaurus et Tyrannosaurus sont des taxons frères, voire le même genre, et Daspletosaurus un parent plus ancestral[27],[29]. D'un autre côté, Philip John Currie et des collègues ont trouvé que Daspletosaurus était plus apparenté au Tarbosaurus et à d'autres tyrannosauridés asiatiques comme Alioramus, plutôt qu'au Tyrannosaurus nord-américain[30]. La systématique de Daspletosaurus sera surement plus claire une fois que les espèces seront toutes décrites.
Lythronax
Lythronax (« roi du sang », formé des mots grecs lythron, sang, et anax, roi) est l'un des genres de dinosaure Theropoda de la famille des Tyrannosauridae. Il a vécu il y a quelque 80 millions d'années dans ce qui est maintenant le sud de l'Utah. Lytronax argestes (l'épithète fait allusion à la localisation du fossile découvert, le sud-ouest de l'Amérique[34]) est, d'après sa position stratigraphique, le plus ancien des Tyrannosauridae connu[35].
Paléobiologie
Alimentation
Selon Irmis, les dents de L. argentes n'étaient pas tout à fait aussi grosses que celles de T. rex, mais étaient comme des couperets à viande en forme de banane, dentés comme un couteau-scie, solides. Elles servaient non seulement à couper la chair, mais aussi à infliger des dommages et à écraser des os[36].
Posture

Comme beaucoup de dinosaures bipèdes, Tyrannosaurus rex a été, de façon erronée, historiquement décrit comme un tripode marchant sur ses deux pattes postérieures avec sa queue servant d'appui au sol : avec le corps presque à la verticale, un peu à la façon d'un kangourou. Cette conception date de la reconstitution en 1865 d'un Hadrosaurus par le paléontologue américain Joseph Leidy, qui fut le premier à décrire un dinosaure en bipédie[37].
_2012-05-11_04.jpg)
Henry Fairfield Osborn, ancien président de l'American Museum of Natural History (AMNH) de New York, pensant que l'animal se tenait à la verticale, inaugura la première reconstitution d'un squelette complet de T. rex en 1915. Celui-ci resta en position debout pendant près d'un siècle, jusqu'à son démantèlement en 1992[38].
Vers 1970, les scientifiques réalisent que cette posture en position verticale avec trois points d'appui n'est pas anatomiquement possible ; elle aurait entraîné la luxation ou la détérioration de nombreuses articulations telles que celles des hanches ou celle comprise entre le crâne et les premières vertèbres cervicales[39].
La reconstitution fausse à l'AMNH a inspiré de nombreux films et peintures jusque dans les années 1990, lorsque les films comme Jurassic Park ont représenté le tyrannosaure dans une posture plus réaliste. Les représentations modernes dans les musées, l'art et les films montrent le Tyrannosaurus rex avec le corps presque parallèle au sol et la queue tendue à l'arrière pour équilibrer la tête.
Bras

Lorsqu'on a découvert le Tyrannosaurus rex, l'humérus était la seule partie connue des pattes antérieures[40]. Le premier squelette montré au public en 1915 et monté par Osborn montraient le Tyrannosaurus rex avec une main à trois doigts, comme pour l'Allosaurus[41]. Pourtant un an plus tôt, Lawrence Lambe avait décrit les membres antérieurs de Gorgosaurus, une espèce proche et apparentée au T. rex, comme étant courts et munis de deux doigts seulement[42]. Ce qui pouvait suggérer que Tyrannosaurus rex avait des bras similaires, hypothèse qui ne fut confirmée que bien plus tard, lorsque les premiers restes de bras entiers furent identifiés en 1989, appartenant à MOR 555, surnommé le "Wankel rex", découvert à Hell Creek[43]. Les restes de "Sue" comprennent également des bras complets[17].

Les bras du Tyrannosaurus rex sont très petits par rapport au reste du corps, mesurant seulement 1 mètre de long. Cependant ces bras ne sont pas des structures vestigiales et montrent de larges zones d'attachement musculaires, indiquant une force considérable. Dès 1906, Osborn avait imaginé que les bras pouvaient être utilisés pour tenir fortement le partenaire lors de l'accouplement[44]. Il a été également suggéré que les bras soient utilisés pour aider l'animal à se redresser à partir d'une position couchée[39]. Ou pour tenir sa proie alors que les mâchoires font le reste, hypothèse qui serait soutenue par des analyses biomécaniques. Ainsi les os des bras possèdent une corticale particulièrement résistante. Le muscle biceps brachial d'un adulte était capable de soulever à lui seul une charge de 199 kg, chiffre augmentant avec l'action combinée des autres muscles du bras comme le brachial. Les mouvements des bras étaient limités, avec des amplitudes articulaires de respectivement 40 et 45 ° aux épaules et aux coudes - en comparaison, chez Deinonychus, les amplitudes de ces deux mêmes articulations sont 88 et 130 °, et chez l'homme 360 et 165 °. Ainsi, les bras du Tyrannosaurus rex, avec leur structure osseuse particulièrement solide, la force considérable développée par leurs muscles et leur amplitude de mouvement limitée, pourrait indiquer un système anatomique conçu pour agripper vite et fort une proie se débattant[45].
Peau et plumes

En 2004, la revue scientifique Nature a publié un rapport décrivant un tyrannosauridé, le Dilong paradoxus, un proche parent du T. Rex apparu avant ce dernier, retrouvé sur le site géologique de la formation Jehol en Chine. Comme de nombreux théropodes découverts dans cette formation, le squelette fossilisé a été préservé dans un manteau de structures filamenteuses reconnues comme étant des structures précurseurs des plumes. C'est ainsi qu'on a également supposé que le Tyrannosaurus et d'autres tyrannosauridés proches possédaient des protoplumes. Cependant, des empreintes de peau retrouvées sur de grands tyrannosauridés montrent des écailles en mosaïques[46]. Il est possible que ces protoplumes aient existé sur des parties du corps qui n'ont pas été préservé. Mais les gros animaux ont, proportionnellement à leur volume, une surface moins importante que les plus petits. Donc plus un animal est gros, moins il souffre des déperditions de chaleur. Les protoplumes deviendraient alors inutiles et pourraient avoir été secondairement perdues lors de l'évolution des grands tyrannosauridés comme Tyrannosaurus, particulièrement sous le chaud climat du crétacé[47]. En 2012 un Tyrannosauroidea basal de très grande taille (les spécimens adultes atteignaient 9 mètres de long pour 1 414 kg) du nom de Yutyrannus fut découvert dans la province du Liaoning avec des traces de plumes sur le cou, la fin de la queue, les jambes et les bras, ce qui suggère que sous les climats froids (des températures annuelles de 10 °C pour la province de Liaoning il y a 125 millions d'années), les Tyrannosaures portaient des plumes qui servaient d'isolant.
Thermorégulation
On a longtemps pensé que le Tyrannosaurus, comme la plupart des dinosaures était poïkilotherme, c'est-à-dire qu'il ne contrôlait pas sa température corporelle et avait le « sang froid », tout comme les reptiles. C'est dans les années 1960 que des scientifiques comme Robert T. Bakker et John Ostrom ont émis l'idée que le métabolisme des dinosaures ressemblait davantage à celui des mammifères et des oiseaux qu'à celui des animaux à sang froid[48],[49]. À la suite de l'analyse d'un squelette, des scientifiques ont déclaré que le Tyrannosaurus rex était homéotherme (à sang chaud), impliquant ainsi une vie très active[50]. Depuis, plusieurs paléontologues ont cherché à déterminer la capacité du Tyrannosaurus à réguler la température de son corps. Des preuves histologiques de taux de croissance rapides chez le jeune T. rex, comparables à ceux des mammifères et des oiseaux, pourrait supporter l'hypothèse d'un métabolisme élevé. Les courbes de croissance indiquent que, tout comme chez les mammifères et les oiseaux, la croissance du Tyrannosaurus rex est limitée à l'âge adulte, contrairement à la croissance indéterminée retrouvée chez de nombreux autres vertébrés[3].
Les ratios d'isotopes d'oxygène présents dans les os fossilisés sont utilisés pour déterminer la température à laquelle l'os a été formé, car ces ratios sont corrélés avec la température. Chez un spécimen de T. rex on a retrouvé des ratios indiquant que la différence de température entre une vertèbre du thorax et un tibia n'était que de 4 à 5 °C. Selon le paléontologue Reese Barrick et le géochimiste William Showers cette différence minime entre le corps de l'animal et ses extrémités indique que Tyrannosaurus rex maintenait sa température interne corporelle constante, définition de l'homéothermie, et qu'il possédait un métabolisme situé entre celui des reptiles ectothermes et des mammifères endothermes[51]. D'autres scientifiques ont fait remarquer que le ratio des isotopes d'oxygène présent dans les fossiles actuels était probablement différent du ratio originel au temps où Tyrannosaurus vivait, et qu'il avait dû être modifié durant ou après le processus de fossilisation appelé la diagenèse[52]. Barrick et Showers se sont défendus en retrouvant des résultats similaires chez un autre dinosaure théropode, le Giganotosaurus, vivant sur un autre continent et dix millions d'années plus tôt[53].
Des ornithischiens montrent également des preuves d'homéothermie, alors que ce n'est pas le cas pour des varans retrouvés dans la même formation[54]. Même si le Tyrannosaurus rex montre des preuves d'homéothermie, cela ne signifie pas obligatoirement qu'il était endotherme. Une telle thermorégulation peut s'expliquer par la gigantothermie, comme chez certaines tortues de mer[55],[56]. Ainsi certains grands poïkilothermes, grâce à un rapport volume / surface de peau favorable, sont capables de maintenir une température de corps et un métabolisme relativement élevés.
Locomotion

Il y a deux questions principales qui font débat concernant les capacités locomotrices du Tyrannosaurus : comment pouvait-il tourner et quelle était sa vitesse de pointe. Ces deux questions sont liées au débat concernant le fait de savoir s'il était chasseur ou charognard.
Le Tyrannosaurus devait être lent à se retourner, prenant probablement une à deux secondes pour se tourner de 45° — à titre de comparaison l'Homme, qui se tient debout et n'a pas de queue, peut se retourner en une fraction de seconde[57]. Cette lenteur est due au moment d'inertie, qui quantifie la résistance d'un corps soumis à une mise en rotation et qui est particulièrement important chez le Tyrannosaurus, dont une grande partie de la masse est située à distance de son centre de gravité. Il pouvait diminuer cette résistance en arquant son dos et sa queue et en rapprochant sa tête et ses bras de son corps, à la façon d'un patineur qui se regroupe pour tourner sur lui-même plus vite[58].
Les scientifiques ont avancé de nombreux chiffres concernant la vitesse maximale de course de Tyrannosaurus rex, la plupart autour de 11 m/s soit 40 km/h, avec des minima de 5–11 m/s (18–40 km/h) et des maxima autour de 20 m/s (72 km/h). Diverses techniques d'estimation ont été utilisées pour aboutir à ces chiffres car, s'il existe de nombreuses traces de pas de grands théropodes en train de marcher, aucune trace de théropodes en train de courir n'a été encore retrouvé, ce qui pourrait indiquer qu'ils ne couraient simplement pas[59]. Les scientifiques qui pensent que Tyrannosaurus était capable de courir soulignent que certaines caractéristiques anatomiques permettent à un adulte de ne peser que 4,5 tonnes et que certains animaux tels que les autruches ou les chevaux, possédant de longues jambes flexibles, sont capables d'atteindre de grandes vitesses grâce à des foulées plus lentes mais plus longues. De plus, certains avancent que Tyrannosaurus avait des muscles aux membres inférieurs plus larges que chez n'importe quel animal vivant actuel, ce qui lui aurait permis de courir jusqu'à 40 à 70 kilomètres par heure[60].
Jack Horner et Don Lessem avancèrent en 1993 que le Tyrannosaurus était lent et ne pouvait probablement pas courir (présence d'une phase de suspension en l'air) car le ratio de la longueur de son fémur sur celle de son tibia était supérieur à 1, comme chez la plupart des grands théropodes et comme les éléphants modernes[43]. Cependant, Holtz écrit en 1998 que les tyrannosauridés et des groupes proches avaient les éléments distaux des membres postérieures (cheville, métatarse et orteils) significativement plus longs que la longueur du fémur de la plupart des autres théropodes, et que les tyrannosauridés et des groupes proches avait un métatarse plus efficace pour transmettre les forces de locomotion du pied à la jambe que chez les premiers théropodes. Il conclut que les tyrannosauridés et des groupes proches étaient les plus rapides des larges théropodes[61].
Christiansen écrit en 1998 que les os de la jambe de Tyrannosaurus n'étaient pas significativement plus solides que ceux des éléphants, qui sont relativement limités concernant leur vitesse de pointe et qui n'ont jamais pu courir (pas de phase de suspension en l'air), et il proposa que la vitesse maximale de Tyrannosaurus devait être d'environ 11 mètres par seconde (40 km/h), ce qui est comparable à la vitesse d'un sprinter humain. Il fit remarquer que cette estimation dépendait de nombreuses hypothèses douteuses[62].
Farlow et ses collègues avancent en 1995 qu'un Tyrannosaurus pesant 5,4 à 7,3 tonnes serait gravement voire fatalement blessé s'il tombait alors qu'il était en train de se mouvoir rapidement, car son thorax frapperait le sol avec une décélération de 6 g sans que ses petits bras puissent réduire l'impact[63]. Cependant, des animaux comme les girafes peuvent galoper à 50 km/h malgré le risque que cela représente[64],[65] et il est donc possible que Tyrannosaurus ait également pu se déplacer rapidement en cas de nécessité et malgré les risques que cela pouvait représenter[66],[67].
Des études récentes sur la vitesse de déplacement de Tyrannosaurus avançaient une allure modérée ne dépassant pas 40 km/h. Par exemple, en 2002 dans la revue Nature, un modèle mathématique est présenté[68] dont le but est de permettre d'estimer la masse musculaire nécessaire au niveau des jambes pour courir rapidement, à plus de 40 km/h[60]. L'article conclut que des vitesses supérieures à 40 km/h sont impossibles à atteindre car elles nécessiteraient de très gros muscles représentant plus de 40 à 86 % de la masse corporelle totale. Si ces muscles avaient été moins massifs, seule une vitesse d'environ 18 km/h aurait pu être atteinte. Faire des conclusions sur cette modélisation est difficile car on ne sait pas quel était le volume des muscles de Tyrannosaurus[60].
Une étude de 2007 utilisant des modèles informatiques estimait la vitesse maximale de course de T. rex à 8 mètres par seconde, soit 29 km/h[69], soit légèrement plus vite qu'un footballeur professionnel et moins vite qu'un sprinter qui peut atteindre 12 mètres par seconde soit 43 km/h[70].
Ceux qui estiment que Tyrannosaurus était incapable de courir estiment sa vitesse de pointe à 17 km/h, ce qui est toujours plus rapide que les hadrosauridés et les cératopsiens qui devaient être ses principales proies[60]. De plus, certains de ceux qui défendent le fait que Tyrannosaurus était un prédateur avancent que sa vitesse de course n'était pas si cruciale, car même s'il était lent, il était toujours plus rapide que ses proies[71]. Cependant, Paul et Christiansen (2000) écrivent que les derniers Ceratopsia avaient des pattes arrières verticales et que les gros spécimens devaient pouvoir courir aussi vite que les rhinocéros[72]. On a retrouvé des fossiles de Ceratopsia présentant des cicatrices de morsures de Tyrannosaurus. Or, si les Ceratopsia ayant vécu en même temps que les T. rex étaient rapides, cela jette le doute sur l'argument que Tyrannosaurus n'avait pas à être rapide pour attraper ses proies[67].
Empreintes de pas

Deux différentes empreintes de pas fossilisées ont été proposées comme appartenant à Tyrannosaurus rex. La première a été découverte au Philmont Scout Ranch dans l'État du Nouveau-Mexique, en 1983, par le géologue américain Charles Pillmore. On pensait initialement qu'elle appartenait à un hadrosauridé ou « dinosaure à bec de canard » mais l'examen attentif de l'empreinte de pas révéla d'une part un large talon inconnu chez les ornithopodes et d'autre part les restes de ce qui aurait pu être un hallux, quatrième orteil en forme d'ergot. Cette empreinte fut décrite comme appartenant à l'ichnotaxon Tyrannosauripus pillmorei en 1994 par en:Martin Lockley et Adrian Hunt, qui suggéraient qu'elle pouvait avoir été faite par un Tyrannosaurus rex, ce qui en ferait la première empreinte de pas connue de l'espèce. La trace est imprimée dans ce qui fut autrefois de la boue provenant d'une terre humide végétale. Elle mesure 83 cm de longueur sur 71 cm de largeur[73].
Une seconde empreinte de pas qui pourrait avoir été faite par un Tyrannosaurus fut découverte dans la formation de Hell Creek dans le Montana en 2007 par le paléontologue britannique Phil Manning. Cette seconde trace de 76 cm de long est plus petite que celle décrite par Lockley et Hunt. Qu'elle appartienne à un Tyrannosaurus n'est pas certain, bien que Tyrannosaurus et Nanotyrannus sont les deux seuls grands théropodes qui ont été retrouvés à Hell Creek. Des études plus approfondies, comparant notamment cette trace à celle du Nouveau-Mexique, sont prévues[74].
Alimentation
Le débat concernant le comportement charognard ou prédateur du Tyrannosaurus est aussi vieux que celui sur sa locomotion.
En 1917, Lambe étudie un squelette de Gorgosaurus, une espèce proche de Tyrannosaurus, et conclut que le Tyrannosaurus était un pur charognard car les dents du Gorgosaurus ne montraient aucune trace d'usure[75]. Cet argument n'est plus considéré comme valide depuis que l'on sait que les théropodes remplacent leurs dents rapidement.
Depuis la première découverte de Tyrannosaurus, la plupart des scientifiques ont la certitude qu'il est un prédateur, bien que tout comme de nombreux prédateurs modernes, il aurait pu aussi voler la prise d'un autre prédateur ou être parfois charognard si l'occasion se présentait[76].
Jack Horner est le principal défenseur de la théorie du tyrannosaure charognard[43],[77],[78]. Il a présenté plusieurs arguments défendant cette hypothèse :
- Les bras du tyrannosaure sont courts comparés à ceux des prédateurs connus. Selon Horner, les bras sont trop courts pour avoir la force nécessaire pour maintenir une proie[79].
- Les tyrannosaures avaient de grands bulbes et nerfs olfactifs par rapport à la taille de leur cerveau. Cela suppose qu'ils possédaient un odorat bien développé qui aurait pu leur permettre de repérer des carcasses à de grandes distances comme les vautours modernes. Des recherches faites sur les bulbes olfactifs de vingt et un dinosaures démontrent que c'est le tyrannosaure qui avait l'odorat le plus développé[80].
- Les dents du tyrannosaure pouvaient broyer les os, et donc extraire des carcasses, généralement les parties les moins nutritives, autant de nourriture (la moelle osseuse) que possible. Karen Chin et son équipe ont trouvé des fragments d'os dans des coprolithes (excréments fossilisés) attribués à des tyrannosaures, mais ils indiquent que les dents du tyrannosaure n'étaient pas adaptées pour broyer systématiquement des os comme le font les hyènes pour extraire la moelle[81].
- Comme les proies potentielles du Tyrannosaurus pouvaient se déplacer rapidement, les preuves qu'il ne pouvait pas courir vite démontrent qu'il était un charognard[77],[82]. D'un autre côté, des analyses récentes démontrent que le Tyrannosaurus bien que plus lent que certains prédateurs terrestres modernes pouvait être assez rapide pour chasser de grands hadrosaures et des cératopsiens[60],[71].

D'autres preuves suggèrent un comportement de prédateur. Les orbites du tyrannosaure étaient placées vers l'avant, lui donnant ainsi une vision binoculaire ce qui lui permettait d'évaluer les distances, bien mieux que les faucons modernes. Horner note aussi une amélioration importante de la vision binoculaire tout au long de l'évolution des tyrannosauridés, avançant qu'il n'était pas évident que la sélection naturelle favorise cette caractéristique si les tyrannosaures avaient été de purs charognards, pour qui une vision stéréoscopique n'aurait pas été un atout[83],[84]. Chez les animaux modernes, la vision binoculaire est principalement retrouvée chez les prédateurs[10].
Selon certains scientifiques, si le Tyrannosaurus était un pur charognard, un autre dinosaure devait être le superprédateur du Crétacé supérieur amérasien. Les autres tyrannosauridés partageant les caractéristiques de Tyrannosaurus, seuls les petits dromæosauridés auraient le potentiel pour être ce superprédateur. Les proies en haut de la chaîne alimentaire étaient alors les marginocéphales et les ornithopodes. Des supporters de l'hypothèse du charognard suggèrent que la taille et la puissance de Tyrannosaurus lui aurait alors permis de voler les proies tuées par de plus petits prédateurs[82]. La plupart des paléontologues acceptent l'hypothèse que le Tyrannosaurus était à la fois prédateur et charognard, comme beaucoup de grands carnivores.
D'autres paléontologues suggèrent que les Tyrannosaures vivaient en groupes familiaux et chassaient en interaction. Les plus jeunes, plus petits, légers et habiles, auraient rabattu les proies vers les adultes embusqués. Ce type de comportement aurait peut-être été adopté par d'autres dinosaures carnivores de grandes tailles comme les allosaures[réf. nécessaire].
Salive infectée
William Abler (en) a proposé que Tyrannosaurus avait peut-être une salive infectieuse, qu'il utilisait pour tuer sa proie[85]. En examinant les dents de tyrannosaures, il s'est aperçu qu'il existait des dentelures pouvant, comme chez le Dragon de Komodo, retenir des morceaux de carcasse permettant la prolifération de bactéries, rendant leur morsure infectante. Horner note cependant que les dentelures de Tyrannosaurus ressemblent plus à des cubes alors que celles des Komodo sont arrondies[86].
Description
Il est connu par un maxillaire et un dentaire associé (os de la mâchoire inférieure portant des dents) récupérés dans le groupe de Wangshi, datant d'environ 70 millions d'années. C'est l'un des plus grands théropodes carnivores connus, mesurant environ 11 m de long, 4 mètres de haut et pesant aux alentours de 6 tonnes. Son dentaire est seulement un peu plus petit que la plus grande mâchoire de Tyrannosaurus rex retrouvé[87],[88].
Il a été trouvé dans ce qui était une plaine inondable au Crétacé et qui contient l'une des plus fortes quantités d'os de dinosaures au monde[89].
Dans la culture populaire
les Tyrannosaurinae sont principalements connus graçe à Tyrannosaurus rex qui apparait dans de nombreux films, livres, series et jeux vidéo.
Annexes
Articles connexes
Références taxinomiques
- Référence Fossilworks Paleobiology Database : Nanotyrannus, Bakker et al. (1988) (en)
- Référence Fossilworks Paleobiology Database : Nanotyrannus lancensis, Gilmore (1946) (en)
Bibliographie
- Tyrannosaurus rex, the tyrant king. Par Pedro Larson, Peter L. Larson, Kenneth Carpenter. Édition: illustrated. Publié par Indiana University Press, 2008. (ISBN 0-253-35087-5), 9780253350879. 435 pages. Aperçu en ligne
Liens externes
- (fr) Description du dinosaure Tyrannosaurus sur PaleoWiki
- (fr) Tyrannosaurus sur Gigadino
- (fr) Le tyrannosaure sur Terra Nova
Notes et références
- ↑ (fr) « Le tyrannosaure Rex souffrait-il de goutte? », sur European Vegetarian Union,
- 1 2 3 (en) GM Erickson, Gregory M., « Gigantism and comparative life-history parameters of tyrannosaurid dinosaurs », Nature, vol. 430, no 7001, , p. 772–775 (PMID 15306807, DOI 10.1038/nature02699)
- 1 2 (en) JR Horner et K Padian, « Age and growth dynamics of Tyrannosaurus rex », Proceedings. Biological sciences / the Royal Society, vol. 271, no 1551, , p. 1875–80 (lire en ligne)
- ↑ (en) Andrew H. Lee et Sarah Werning, « Sexual maturity in growing dinosaurs does not fit reptilian growth models », Proceedings of the National Academy of Sciences, vol. 105, no 2, , p. 582–587 (DOI 10.1073/pnas.0708903105)
- ↑ (en) Andrew H. Lee et Sarah Werning, « Sexual maturity in growing dinosaurs does not fit reptilian growth models », Proc Natl Acad Sci U S A, vol. 105, no 2, 2008 jan 15, p. 582-7 (lire en ligne)
- 1 2 « Maturité sexuelle précoce chez les dinosaures », sur http://www.radio-canada.ca, (consulté le 17 mars 2009)
- 1 2 (en) MH Schweitzer, JL Wittmeyer et JR Horner, « Gender-specific reproductive tissue in ratites and Tyrannosaurus rex », Science (New York, N.Y.), vol. 308, no 5727, , p. 1456–60 (DOI 10.1126/science.1112158, lire en ligne)
- ↑ « Rapid Growth, Early Maturity Meant Teen Pregnancy For Dinosaurs », sur http://www.sciencedaily.com, ScienceDaily, (consulté le 17 mars 2009)
- 1 2 3 (en) GM Erickson, PJ Currie, BD Inouye et AA Winn, « Tyrannosaur life tables: an example of nonavian dinosaur population biology », Science (New York, N.Y.), vol. 313, no 5784, , p. 213–7 (DOI 10.1126/science.1125721, lire en ligne)
- 1 2 (en) « New Dinosaur Species, Lythronax Argestes, Discovered In Utah », sur LiveScience, Stephanie Pappas (consulté le 8 novembre 2013).
- ↑ « Lythronax, l'ancêtre du redoutable T. rex », sur L'Orient-Le Jour.com, (consulté le 9 novembre 2013).
- ↑ (en) Kenneth Carpenter, Dinosaur Systematics: Approaches and Perspectives, Cambridge, Kenneth Carpenter et Philip John Currie, (réimpr. Cambridge University Press), 141–145 p. (ISBN 0-521-43810-1), « Variation in Tyrannosaurus rex »
- ↑ Larson, P.L. 1994. Tyrannosaurus sex. In: Rosenberg, G.D. & Wolberg, D.L. Dino Fest. The Paleontological Society Special Publications. 7: 139–155.
- ↑ Romer, A. S. 1956. Osteology of the Reptiles. University of Chicago, Chicago, Illinois, 800 p.
- ↑ (en) Erickson GM, Kristopher Lappin A, Larson P, « Androgynous rex – The utility of chevrons for determining the sex of crocodilians and non-avian dinosaurs », Zoology (Jena), vol. 108, no 4, , p. 277-86 (résumé)
- ↑ « All about Sue - Important Discoveries », sur http://www.aucklandmuseum.com, (consulté le 13 avril 2009)
- 1 2
- ↑ Schweitzer MH, Elsey RM, Dacke CG, Horner JR, Lamm ET, « Do egg-laying crocodilian (Alligator mississippiensis) archosaurs form medullary bone? », Bone, vol. 40, no 4, , p. 1152–8 (PMID 17223615, DOI 10.1016/j.bone.2006.10.029, lire en ligne)
- ↑ (en) Darren H. Tanke, « Head-biting behavior in theropod dinosaurs: paleopathological evidence », Gaia, vol. 15, , p. 167–184 (lire en ligne [PDF]) [not printed until 2000]
- ↑
- ↑ (en) David A. Eberth, « Were large theropods gregarious? », Journal of Vertebrate Paleontology, vol. 21, no (Supplement to 3 - Abstracts of Papers, 61st Annual Meeting of the Society of Vertebrate Paleontology), , p. 46A[published abstract only]
- ↑ (en) Brian T. Roach, « A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs », Bulletin of the Peabody Museum of Natural History, vol. 48, no 1, , p. 103–138 (DOI [103:AROCPH2.0.CO;2 10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2], lire en ligne)
- 1 2 (en) James O. Farlow, « Body size overlap, habitat partitioning and living space requirements of terrestrial vertebrate predators: implications for the paleoecology of large theropod dinosaurs », Historical Biology, vol. 16, no 1, , p. 21–40 (DOI 10.1080/0891296031000154687)
- ↑ (en) Dale A. Russell, « Tyrannosaurs from the Late Cretaceous of western Canada », National Museum of Natural Sciences Publications in Paleontology, vol. 1, , p. 1–34
- ↑
- ↑ (en) Eric Snively, « Fused and vaulted nasals of tyrannosaurid dinosaurs: implications for cranial strength and feeding mechanics », Acta Palaeontologica Polonica, vol. 51, no 3, , p. 435–454 (lire en ligne [PDF])
- 1 2 3 (en) Thomas R. Holtz, The Dinosauria, Berkeley, University of California Press, , 111–136 p. (ISBN 0-520-24209-2), « Tyrannosauroidea »
- ↑ « A new study of the brain of the predatory dinosaur Tarbosaurus bataar », Paleontological Journal, vol. 41, no 3, , p. 281-289 (DOI 10.1134/S0031030107030070, lire en ligne)
- 1 2 (en) Thomas D. Carr, « A new genus and species of tyrannosauroid from the Late Cretaceous (middle Campanian) Demopolis Formation of Alabama », Journal of Vertebrate Paleontology, vol. 25, no 1, , p. 119–143 (DOI [0119:ANGASO2.0.CO;2 10.1671/0272-4634(2005)025[0119:ANGASO]2.0.CO;2], lire en ligne)
- 1 2 (en) Philip J. Currie, « Skull structure and evolution in tyrannosaurid phylogeny », Acta Palaeontologica Polonica, vol. 48, no 2, , p. 227–234 (lire en ligne [PDF])
- ↑ (en) John R. Horner, « Marine transgressions and the evolution of Cretaceous dinosaurs », Nature, vol. 358, no 6381, , p. 59–61 (DOI 10.1038/358059a0)
- ↑ (en) Gregory S. Paul, Predatory Dinosaurs of the World, New York, Simon & Schuster, , 464pp p. (ISBN 978-0-671-61946-6)
- ↑ (en) Thomas D. Carr, « Craniofacial ontogeny in Tyrannosauridae (Dinosauria, Coelurosauria) », Journal of Vertebrate Paleontology, vol. 19, no 3, , p. 497–520 (DOI 10.1080/02724634.1999.10011161)
- ↑ Maxime Lambert, « Le Lythronax, le plus vieil ancêtre du Tyrannosaurus rex jamais découvert », sur maxisciences.com, (consulté le 9 novembre 2013).
- ↑ (en) M. A. Loewen, R. B. Irmis, J. J. W. Sertich, P. J. Currie et S. D. Sampson, « Tyrant Dinosaur Evolution Tracks the Rise and Fall of Late Cretaceous Océans », PLoS One, vol. 8, no 11, , e79420 (DOI 10.1371/journal.pone.0079420).
- ↑ (en) « King of gore dinosaur was the 'bad grandpa' of tyrannosaurs », sur Los Angeles Times, Geoffrey Mohan (consulté le 8 novembre 2013).
- ↑ (en) Joseph Leidy, « Memoir on the extinct reptiles of the Cretaceous formations of the United States », Smithsonian Contributions to Knowledge, vol. 14, , p. 1–135
- ↑ « Tyrannosaurus », American Museum of Natural History (consulté le 15 avril 2009)
- 1 2 BH Newman, « Stance and gait in the flesh-eating Tyrannosaurus », Biological Journal of the Linnean Society, vol. 2, , p. 119–123 (résumé)
- ↑ (en) H. F. Osborn, « Tyrannosaurus and other Cretaceous carnivorous dinosaurs », Bulletin of the AMNH, New York, American Museum of Natural History, vol. 21, no 14, , p. 259–265 (lire en ligne)
- ↑ (en) H. F. Osborn, « Skeletal adaptations of Ornitholestes, Struthiomimus, Tyrannosaurus », Bulletin of the American Museum of Natural History, New York, American Museum of Natural History, vol. 35, no 43, , p. 733–771 (lire en ligne)
- ↑ (en) L. M. Lambe, « On a new genus and species of carnivorous dinosaur from the Belly River Formation of Alberta, with a description of the skull of Stephanosaurus marginatus from the same horizon », Ottawa Naturalist, vol. 27, , p. 129–135
- 1 2 3 (en) John R. Horner, Don Lessem, The complete T. rex, New York, Simon & Schuster, (ISBN 978-0-671-74185-3, LCCN 93000211)
- ↑ (en) Henry Fairfield Osborn, « Tyrannosaurus, Upper Cretaceous carnivorous dinosaur », Bulletin of the AMNH, New York, American Museum of Natural History, vol. 22, no 16, , p. 281–296 (lire en ligne)
- ↑ (en) Kenneth Carpenter et Matt Smith, « Forelimb Osteology and Biomechanics of Tyrannosaurus rex », dans Darren H. Tanke & Kenneth Carpenter (éditeurs) et Michael W. Skrepnick (directeur artistique), Mesozoic vertebrate life, Bloomington, Indiana University Press, (ISBN 0-253-33907-3, lire en ligne)
- ↑ (en) Gregory S. Paul, Tyrannosaurus rex, the Tyrant King (Life of the Past), Bloomington, Indiana University Press, (ISBN 978-0-253-35087-9), « The extreme lifestyles and habits of the gigantic tyrannosaurid superpredators of the Late Cretaceous of North America and Asia », p. 316
- ↑ (en) « Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids », Nature, vol. 431, no 7009, , p. 680–684 (PMID 15470426, DOI 10.1038/nature02855)
- ↑ (en) Robert T. Bakker, « The superiority of dinosaurs », Discovery, vol. 3, no 2, , p. 11–12 (lire en ligne [PDF])
- ↑ (en) Robert T. Bakker, « Anatomical and ecological evidence of endothermy in dinosaurs », Nature, vol. 238, , p. 81–85 (DOI 10.1038/238081a0, lire en ligne [PDF])
- ↑ Robert T. Bakker, The Dinosaur Heresies, New York, Kensington Publishing, (ISBN 0-688-04287-2, OCLC 13699558)
- ↑ (en) Reese E. Barrick, « Thermophysiology of Tyrannosaurus rex: Evidence from Oxygen Isotopes », Science, New York, vol. 265, no 5169, , p. 222–224 (PMID 17750663, DOI 10.1126/science.265.5169.222, lire en ligne)
- ↑ (en) Clive Trueman, « Diagenetic effects on the oxygen isotope composition of bones of dinosaurs and other vertebrates recovered from terrestrial and marine sediments », Journal of the Geological Society, vol. 160, no 6, , p. 895 (DOI 10.1144/0016-764903-019)
- ↑ (en) Reese E. Barrick, « Thermophysiology and biology of Giganotosaurus: comparison with Tyrannosaurus », Palaeontologia Electronica, vol. 2, no 2, (lire en ligne)
- ↑ (en) Reese E. James O. Farlow (éditeur) et M. K. Brett-Surman (éditeur), Michael K. Stoskopf et William J. Showers, The Complete Dinosaur, Bloomington, Indiana University Press, (ISBN 0-253-21313-4), « Oxygen isotopes in dinosaur bones »
- ↑ (en) Frank V. James O. Farlow (éditeur) et M. K. Brett-Surman (éditeur), James R. Spotila et Peter Dodson, The Complete Dinosaur, Bloomington, Indiana University Press, (ISBN 0-253-21313-4), « A blueprint for giants: modeling the physiology of large dinosaurs »
- ↑ (en) Anusuya David B. Weishampel (éditeur), Peter Dodson (éditeur) et Halszka Osmólska (éditeur), Willem J. Hillenius, The dinosauria, Berkeley, University of California Press, (ISBN 0-520-24209-2), « Physiology of nonavian dinosaurs »
- ↑ JR Hutchinson JR, V Ng-Thow-Hing V et FC Anderson, « A 3D interactive method for estimating body segmental parameters in animals: application to the turning and running performance of Tyrannosaurus rex », Journal of theoretical biology, vol. 246, no 4, , p. 660–80 (DOI 10.1016/j.jtbi.2007.01.023)
- ↑ (en) David R. Carrier, « Influence of rotational inertia on turning performance of theropod dinosaurs: clues from humans with increased rotational inertia », Journal of Experimental Biology (en), the Company of Biologists (en), vol. 204, no 22, , p. 3917–3926 (PMID 11807109, lire en ligne)
- ↑ (en) Hutchinson, J.R., « Biomechanical Modeling and Sensitivity Analysis of Bipedal Running Ability. II. Extinct Taxa », Journal of Morphology, vol. 262, no 1, , p. 441–461 (PMID 15352202, DOI 10.1002/jmor.10240)
- 1 2 3 4 5 (en) Hutchinson JR, Garcia M, « Tyrannosaurus was not a fast runner », Nature, vol. 415, no 6875, , p. 1018–21 (PMID 11875567, DOI 10.1038/4151018a)
- ↑ (en) Holtz, « Phylogenetic taxonomy of the Coelurosauria (Dinosauria; Theropoda) », Journal of Paleontology, vol. 70, no 3, , p. 536–538 (lire en ligne)
- ↑ (en) Christiansen, P., « Strength indicator values of theropod long bones, with comments on limb proportions and cursorial potential », Gaia, vol. 15, , p. 241–255 (ISSN 0871-5424, lire en ligne [PDF])
- ↑ (en) JO Farlow, MB Smith et JM Robinson, « Body mass, bone "strength indicator", and cursorial potential of Tyrannosaurus rex », Journal of Vertebrate Paleontology, vol. 15, no 4, , p. 713–725 (lire en ligne)
- ↑ (en) « Giraffe », WildlifeSafari.info (consulté le 29 avril 2006)
- ↑ (en) « The History of Woodland Park Zoo – Chapter 4 » (consulté le 29 avril 2006)
- ↑ (en) Alexander, R.M., « Dinosaur biomechanics », Proc Biol Sci., The Royal Society, vol. 273, no 1596, , p. 1849–1855 (PMID 16822743, PMCID 1634776, DOI 10.1098/rspb.2006.3532)
- 1 2 (en) Rebecca R. Hanna, « Multiple injury and infection in a sub-adult theropod dinosaur (Allosaurus fragilis) with comparisons to allosaur pathology in the Cleveland-Lloyd dinosaur quarry collection », Journal of Vertebrate Paleontology, vol. 22, no 1, , p. 76–90 (DOI [0076:MIAIIA2.0.CO;2 10.1671/0272-4634(2002)022[0076:MIAIIA]2.0.CO;2]) catalogs the injuries of the Allosaurus known as "Big Al" – at least one was attributed to a fall.
- ↑ (en) Hutchinson, J.R., « Biomechanical Modeling and Sensitivity Analysis of Bipedal Running Ability. II. Extinct Taxa », Journal of Morphology, vol. 262, no 1, , p. 441–461 (PMID 15352202, DOI 10.1002/jmor.10240, lire en ligne [PDF])
- ↑ (en) Sellers, W.I., and Manning, P.L., « Estimating dinosaur maximum running speeds using evolutionary robotics », Proc. R. Soc. B, The Royal Society, vol. 274, no 1626, , p. 2711 (PMID 17711833, PMCID 2279215, DOI 10.1098/rspb.2007.0846, lire en ligne)
- ↑ (en) L Seward, « T. rex 'would outrun footballer' », BBCNews, (consulté le 16 octobre 2008)
- 1 2 (en) Manning P (éditeur), Carpenter, Kenneth (éditeur) et Peter E. (éditeur), Tyrannosaurus rex, the Tyrant King (Life of the Past), Bloomington, Indiana University Press, (ISBN 0-253-35087-5), « T. rex speed trap »
- ↑ (en) Paul, G.S. et Christiansen, P., « Forelimb posture in neoceratopsian dinosaurs: implications for gait and locomotion », Paleobiology, vol. 26, no 3, , p. 450 (DOI <0450:FPINDI>2.0.CO;2 10.1666/0094-8373(2000)026<0450:FPINDI>2.0.CO;2, lire en ligne)
- ↑ (en) MG Lockley, « A track of the giant theropod dinosaur Tyrannosaurus from close to the Cretaceous/Tertiary boundary, northern New Mexico », Ichnos, vol. 3, , p. 213–218 (DOI 10.1080/10420949409386390)
- ↑ (en) « T.rex footprint discovered », Natural History Museum, (consulté le 9 décembre 2008)
- ↑ (en) Lambe, L. B., « The Cretaceous theropodous dinosaur Gorgosaurus », Memoirs of the Geological Survey of Canada, vol. 100, , p. 1–84
- ↑ Farlow, J. O. and Holtz, T. R., « The Fossil Record of Predation » [PDF], The Paleontological Society Papers, Kowalewski, M. and Kelley, P.H., , p. 251–266
- 1 2 (en) Horner, J.R., « Steak knives, beady eyes, and tiny little arms (a portrait of Tyrannosaurus as a scavenger) », The Paleontological Society Special Publication, vol. 7, , p. 157–164
- ↑ (en) Amos, J., « T. rex goes on trial », BBC,
- ↑ The king of killers or fearsome freeloader? It is one of the big questions currently in palaeontology.
- ↑ (en) « T. Rex brain study reveals a refined "nose" », Calgary Herald, (consulté le 29 octobre 2008)
- ↑ (en) Chin, K., Erickson, G.M. et al., « A king-sized theropod coprolite », Nature, vol. 393, , p. 680 (DOI 10.1038/31461, lire en ligne) Summary at (en) Monastersky, R., « Getting the scoop from the poop of T. rex », Science News, vol. 153, no 25, , p. 391 (DOI 10.2307/4010364, JSTOR 4010364, lire en ligne)
- 1 2 (en) Walters, Martin, Bloomsbury Illustrated Dictionary of Prehistoric Life (Bloomsbury Illustrated Dictionaries), Godfrey Cave Associates Ltd, (ISBN 978-1-85471-648-4)
- ↑ (en) Kent A. Stevens, « Binocular vision in theropod dinosaurs », Journal of Vertebrate Paleontology, vol. 26, no 2, , p. 321–330 (DOI [321:BVITD2.0.CO;2 10.1671/0272-4634(2006)26[321:BVITD]2.0.CO;2], lire en ligne [PDF])
- ↑ (en) Eric Jaffe, « Sight for 'Saur Eyes: T. rex vision was among nature's best », Science News, vol. 170, no 1, , p. 3 (DOI 10.2307/4017288, JSTOR 4017288, lire en ligne)
- ↑ (en) William Abler, « The Teeth of the Tyrannosaurs », Scientific American, (consulté le 16 mai 2010)
- ↑ John R. Horner, The Complete T. Rex: How Stunning New Discoveries Are Changing our Understanding of the World's Most Famous Dinosaur, 1993, p. 214-215.
- ↑ David W.E. Hone, Kebai Wang, Corwin Sullivan, Xijin Zhao, Shuqing Chen, Dunjin Li, Shuan Ji, Qiang Ji and Xing Xu, « A new tyrannosaurine theropod, Zhuchengtyrannus magnus is named based on a maxilla and dentary », Cretaceous Research, vol. in press, (DOI 10.1016/j.cretres.2011.03.005, lire en ligne)
- ↑ (en) « Giant prehistoric dinosaur cousin of T. rex identified », BBC, (consulté le 3 avril 2011)
- ↑ (en) « New dino in same league as T. rex », Sydney Morning Herald, 1er avril 2011 (consulté le 3 avril 2011)
- Kurzanov, S.M. (1976). [A new Late Cretaceous carnosaur from Nogon-Tsav, Mongolia.] Journal of the Soviet-Mongolian Paleontological Expeditions 3:93-104.
- Holtz, T.R. (2004). Tyrannosauroidea. In: Weishampel, D.A., Dodson, P., and Osmolska, H. (Eds.). The Dinosauria (2nd Edition). University of California Press:Berkeley, 111-136. ISBN 0-520-24209-2
- Currie, P.J., Hurum, J.H., and Sabath, K. 2003. Skull structure and evolution in tyrannosaurid phylogeny. Acta Palaeontologica Polonica 48(2):227-234.
- Xu X., Norell, M.A., Kuang X., Wang. X., Zhao Q., and Jia C. (2004). Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids. Nature 431:680-684.
- Currie, P.J. (2003). Cranial anatomy of tyrannosaurid dinosaurs from the Late Cretaceous of Alberta, Canada. Acta Palaeontologica Polonica 48(2):191–226.
- Carr, T.D., Williamson, T.E., and Schwimmer, D.R. (2005). A new genus and species of tyrannosauroid from the Late Cretaceous (middle Campanian) Demopolis Formation of Alabama. Journal of Vertebrate Paleontology 25(1):119-143.
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Daspletosaurus » (voir la liste des auteurs).
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Deinodon » (voir la liste des auteurs).
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Nanuqsaurus » (voir la liste des auteurs).
 Portail des dinosaures
Portail des dinosaures  Portail de l’herpétologie
Portail de l’herpétologie  Portail de la paléontologie
Portail de la paléontologie