{kind=link}
Productivité primaire
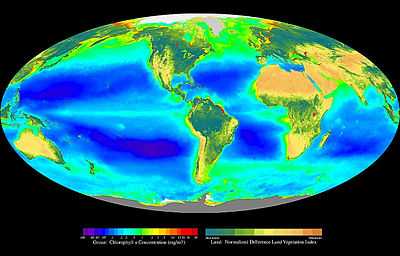
La productivité primaire traduit la vitesse à laquelle se forme par unité de temps, une quantité donnée de matière organique à partir de matière minérale et d'un apport d'énergie. C'est un flux exprimé en masse de carbone assimilé par unité de temps[1]. Cette production de matière organique s'effectue principalement grâce à la photosynthèse (la chimiosynthèse étant beaucoup moins répandue). Dans un écosystème, la productivité primaire est réalisée par les producteurs primaires également appelés autotrophes. Il s'agit du premier maillon d'une chaine alimentaire dans un réseau trophique. Les producteurs primaires sont principalement des plantes dans les écosystèmes terrestres et des algues dans les écosystèmes aquatiques. On appelle production primaire l'énergie qui est accumulée par la productivité primaire. On peut distinguer deux sous-ensembles de production primaire : la Production Primaire Brute et la Production Primaire Nette.
Processus de la productivité primaire
La productivité primaire est l'accumulation d'énergie chimique (la production primaire) sous la forme de matière organique. La source d'énergie utilisée pour réaliser cette production est soit l'énergie lumineuse (dans le cas de la photosynthèse), soit l'énergie issue de l’oxydation de molécules inorganiques telles que les gaz hydrogénés (ex: le sulfure d'hydrogène, H2S) ou le méthane (dans le cas de la chimiosynthèse). Quelle que soit la source d'énergie utilisée, cette source d'énergie sert à synthétiser des molécules organiques complexes à partir de simples composés inorganiques tels que le dioxyde de carbone (CO2) et l'eau (H2O).
Les deux équations ci-dessous sont des représentations simplifiées de la photosynthèse (en haut) et d'une forme de chimiosynthèse (en bas):
- CO2 + H2O + lumière+Sels mineraux
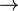 CH2O + O2
CH2O + O2 - CO2 + O2 + 4 H2S CH2O + 4 S + 3 H2O
- CO2 + H2O + lumière+Sels mineraux
Dans tous les cas, ces processus aboutissent à la formation de glucides (CH2O) comme le glucose. Ces molécules relativement simples peuvent ensuite être utilisées pour synthétiser des molécules encore plus complexes comme des protéines, des glucides complexes, des lipides et des acides nucléiques, ou être respirées pour permettre aux cellules de fonctionner (cf. respiration cellulaire). La consommation des producteurs primaires par les organismes hétérotrophes, comme les animaux, transfère ensuite ces molécules organiques (et l'énergie stockée dans ces molécules) vers les autres maillons de la chaîne alimentaire.
Productivité Primaire Brute et Productivité Primaire Nette
- la Production Primaire Brute : énergie totale assimilée par les producteurs primaires (plantes, algues...) par le biais de la photosynthèse ;
- la Production Primaire Nette : quantité d'énergie accumulée dans la biomasse de la plante (croissance et reproduction).
La Production Primaire Nette (PPN) correspond donc à l'énergie accumulée par la plante par le biais de la photosynthèse moins l'énergie utilisée par la plante pour la respiration. En d'autres termes, la différence entre la Production Primaire Brute (PPB) et la Production Primaire Nette correspond à l'énergie utilisée lors de la respiration cellulaire.
- PPN = PPB - respiration
La productivité primaire peut être vue à l'échelle d'une plante (ex: un arbre) ou à celle d'un écosystème.
Dans le cas de l'écosystème, on parle de production primaire nette de l'écosystème.
Cette valeur est utile pour les écologistes dans le sens où elle permet de déterminer la quantité d'énergie disponible pour les autres organismes (détritivores, herbivores et autres maillons de la chaîne alimentaire). Elle est aussi utile pour les activités qui dépendent de la productivité du sol ou d'autres écosystèmes (zones humides, milieux marins, forêts, etc.). La biodiversité est un des facteurs importants de la productivité d'un milieu et de la qualité des services écologiques qu'il rend[2].
Productivité terrestre
Dans les écosystèmes terrestres actuels, la quasi-totalité de la production est effectuée par les plantes vasculaires, avec une faible contribution des algues et des plantes non vasculaires telles que les bryophytes. Dans les temps anciens, avant l'apparition des plantes vasculaires, la contribution des plantes non vasculaires était beaucoup plus importante. La productivité des écosystèmes terrestres dépend de plusieurs facteurs. Les principaux sont la teneur en eau des sols, les précipitations et la température (la température co-varie avec l'énergie solaire). Ces différents facteurs entraînent une répartition différente des végétaux qui permet de différencier les différents biomes terrestres.
Productivité océanique

À l'inverse des écosystèmes terrestres, la productivité des écosystèmes océaniques est principalement assurée par les algues avec une faible contribution des plantes vasculaires. Les algues englobent une grande variété d'organismes allant de simples organismes unicellulaires à des organismes plus complexes comme les macroalgues. Les plantes vasculaires se limitent aux herbiers.
La majorité de la productivité océanique est assurée par le phytoplancton. On estime qu'il est à lui seul responsable de 75 % de la production primaire océanique[3], les autres autotrophes (herbiers et macroalgues) étant souvent limités à des zones restreintes de l'océan : les zones côtières où les eaux sont peu profondes et suffisamment éclairées.
Facteurs de contrôle de la productivité océanique
Les facteurs limitant la production primaire dans les océans sont également très différents de ceux limitant la production primaire des écosystèmes terrestres. La disponibilité en eau n'est de toute évidence plus un problème (bien que sa salinité puisse l'être). De même la température, bien qu'elle puisse affecter les enzymes impliquées dans la photosynthèse, a un rôle limitant beaucoup moins important que dans les écosystèmes terrestres puisque la capacité thermique des eaux limite les variations de température. De plus, l'eau salée gèle à une température plus faible que l'eau douce des écosystèmes terrestres. À l'inverse la lumière, principale source d'énergie de la photosynthèse, et les éléments nutritifs, qui servent à la construction des composants cellulaires et sont utilisés pour la croissance, ont un rôle primordial.
La lumière
La zone éclairée de l'océan est appelée la zone photique (ou de la zone euphotique). Il s'agit d'une couche d'eau relativement mince (0-100 m) près de la surface où il y a suffisamment de lumière pour que la photosynthèse puisse se réaliser. Pour des raisons pratiques, l'épaisseur de la zone photique est généralement définie par la profondeur à laquelle la lumière atteint 1 % de sa valeur en surface. La lumière est atténuée dans la colonne d'eau par son absorption et sa diffraction par l'eau en elle-même, et par les matières dissoutes et particulaires (y compris le phytoplancton).
La capacité à réaliser la photosynthèse dans la colonne d'eau est déterminée par l'interaction entre la zone photique et la couche de mélange. Le mélange turbulent par l'énergie éolienne à la surface de l'océan homogénéise la colonne d'eau à la verticale jusqu'à ce que la turbulence se dissipe (création de ladite couche mélangée). Plus la couche de mélange est importante, plus la quantité moyenne de lumière interceptée par le phytoplancton est importante. La profondeur de la couche de mélange peut varier : elle peut être moins profonde ou plus profonde que la zone photique. Quand elle est beaucoup plus profonde que la zone photique, le phytoplancton passe une grande partie de son temps à l'obscurité ce qui ralentit sa croissance et sa capacité à réaliser la photosynthèse. La profondeur maximale de la couche de mélange où la croissance et la photosynthèse s'effectuent encore de façon efficace est appelée profondeur critique. Tant qu'il y a suffisamment de nutriments disponibles, la production primaire nette se poursuit même lorsque la couche de mélange est moins profonde que la profondeur critique[4].
L'ampleur du brassage des eaux par les vents et la disponibilité de la lumière à la surface des océans sont modifiées à différentes échelles spatiales et temporelles. Les modifications les plus importantes sont liées au cycle des saisons en réponse aux modification de l'angle d'incidence des rayons lumineux à la surface de la Terre. C'est particulièrement vrai dans les écosystèmes tempérés où la productivité primaire est fortement influencée par la lumière incidente à la surface des océans (réduite en hiver) et par l'intensité du mélange de la colonne d'eau plus important pendant la période hivernale en raison des tempêtes. Dans les zones tropicales, la saisonnalité est moins marquée, la lumière incidente étant beaucoup moins variable et le mélange des eaux ne se produisant que de façon épisodique pendant les périodes de tempête.
SHEGUEUY
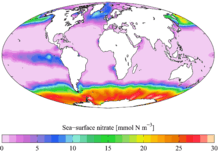
Le mélange des eaux joue également un rôle important dans la limitation de la production primaire par les éléments nutritifs. Les nutriments inorganiques, comme les nitrates, les nitrites, les phosphates et les silicates sont nécessaires au phytoplancton qui les utilisent pour sa croissance et pour le bon fonctionnement de sa machinerie cellulaire. Les composants de l’appareil photosynthétique, les enzymes et les protéines sont, en effet, riches en azote (N) et en fer (Fe). Le phosphore (P) (issu des phosphates) est essentiel à la croissance du phytoplancton et à sa division cellulaire, il est l'un des principaux constituants de diverses biomolécules (tels que les acides nucléiques et les lipides qui sont notamment des composants essentiels des membranes cellulaires). Il est également impliqué dans le transfert de l'énergie et le métabolisme cellulaire (c’est un composant de l’ATP, de l’ADP et du NADP+/NADPH)[5],[6]. Les silicates sont utilisés par certaines diatomées pour la construction de leur squelette externe siliceux. Les cellules phytoplanctoniques sont incapables de synthétiser ces éléments et doivent se les procurer dans leur milieu de vie sous la forme de nutriments inorganiques. Ainsi la limitation de la disponibilité de l’un de ces éléments peut conduire à une diminution de la photosynthèse et par voie de conséquence de la productivité primaire.
Chez le phytoplancton, la limitation en azote (N) peut affecter un certain nombre de processus cellulaires comme la photosynthèse et la synthèse des protéines[7]. La photosynthèse peut être affectée par une réduction de l'efficacité de collecte de l'énergie lumineuse en raison d'une réduction de la quantité de chlorophylle a qui contient de l'azote mais aussi par la réduction de l'efficacité de synthèse des protéines qui constituent l'appareil photosynthétique.
En raison de la chute gravitationnelle de la matière particulaire (comme le plancton, les matières fécales ou les organismes morts), les éléments nutritifs quittent constamment la zone photique, et ne la réalimentent que par le mélange des eaux et la remontée des eaux plus profondes. Cette situation s'aggrave en été en raison du réchauffement des eaux de surface et de la réduction des vents qui augmentent la stratification verticale et conduisent à une forte thermocline, car cela rend plus difficile le mélange des eaux et la remontée des eaux plus profondes riches en éléments nutritifs. Dans les zones d'upwelling, on observe une forte productivité primaire en raison de la remontée importante des eaux profondes riches en éléments nutritifs.
Le Fer
Un autre facteur potentiellement limitant découvert relativement récemment est le fer[8]. Il est utilisé en tant que cofacteur (biochimie) des enzymes impliquées dans les processus tels que la réduction des nitrates et la fixation de l'azote. Une source importante de fer dans les océans provient de la poussière des déserts qui est transportée par les vents. Le Sahara joue ainsi un rôle essentiel dans la fertilisation des océans. Dans les régions de l'océan qui sont éloignées des déserts ou qui ne sont pas atteintes par les vents porteurs de poussières (par exemple, le Sud et le Nord de l'Océan Pacifique), le manque de fer peut limiter considérablement la productivité primaire. Ces zones sont connues sous le nom de zones HNLC pour High-Nutrient, low-chlorophyll (en) en anglais (teneur élevée en nutriments, faible teneur en chlorophylle) en raison de la rareté du fer qui limite la croissance du phytoplancton et conduit à un excès de nutriments. Certains scientifiques ont suggéré d'introduire du fer dans ces régions comme un moyen d'accroître la productivité primaire et l'utilisation du dioxyde de carbone de l'atmosphère afin de limiter les effets du réchauffement climatique[9]. Cependant, des essais de fertilisation par le fer ont montré que l'ajout de fer peut effectivement augmenter la productivité primaire mais que ses effets disparaissent très rapidement. De plus, il est toujours très dangereux de modifier artificiellement la composition des écosystèmes naturels car on ne mesure pas toujours les conséquences que cela pourrait avoir.
Méthodes de mesure
Les méthodes de mesure de la production primaire diffèrent selon le type de production que l'on souhaite mesurer (Brute ou Nette) et en fonction du milieu visé (milieu terrestre ou aquatique). La production brute est toujours plus difficile à mesurer que la production nette car elle nécessite de mesurer la quantité d'énergie vouée à la respiration. La respiration est, en effet, un processus permanent qui consomme une partie des produits de la production primaire (ex : les sucres) avant d'être correctement mesurée. L'étude de la production primaire terrestre est généralement rendu difficile par le fait qu'une partie de cette production est dirigée vers les tissus et organes souterrains plus difficiles d'accès. En milieu aquatique, le même problème se pose avec le phénomène de neige marine et les écosystèmes profonds.
Notes et références
- ↑ Serge Frontier, Denise Pichod-Viale, Alain Leprêtre, Dominique Davoult et Christophe Luczak, Ecosystèmes : Structure, Fonctionnement, Evolution, Dunod, 576 p.
- ↑ Costanza R. et al., 2007, Biodiversity and ecosystem services: A multi-scale empirical study of the relationship between species richness and net primary production
- ↑ (en) Paul G. Falkowski et John A. Raven, Aquatic photosynthesis, Blackwell Publishers, 375 p.
- ↑ Sverdrup, 1953, On condition for the vernal blooming of phytoplankton". Journal du Conseil International pour l'Exploration de la Mer. 18:287-295
- ↑ Frost P.C. and Xenopoulos M.A., 2002, Ambient solar ultraviolet radiation and its effects on phosphorus flux into boreal lake phytoplankton communities". Canadian journal of fisheries and aquatic sciences. 59:1090-1095
- ↑ Iglesias A.A., Plaxton W.C. and Podesta F.E., 1993, The role of inorganic phosphate in the regulation of C4 photosynthesis". Photosynthesis research. 35:205-211
- ↑ Bergers J.A., Charlebois D.A., Mauzerall D.C. and Falkowski P.G., 1996, Differential effects of nitrogen limitation on photosynthetic efficiency of photosystem I and II in microalgae". Plant physiology. 110:689-696
- ↑ (en) J. H. Martin et S. E. Fitzwater, « Iron-deficiency limits phytoplankton growth in the Northeast Pacific Subarctic », Nature, vol. 331, no 6154, , p. 341–343 (DOI 10.1038/331341a0)
- ↑ (en) D.J. Cooper, « Large decrease in ocean—surface Modèle:Co2 fugacity in response to in situ iron fertilization », Nature, vol. 383, no 6600, , p. 511–513 (DOI 10.1038/383511a0)
Annexes
Articles connexes
- Productivité
- Productivité secondaire
- Agriculture
- Agronomie
- Pêche
- Évaluation environnementale
- Élevage
- Plancton
- Phytoplancton
Liens externes
- (fr) Cette section est vide, insuffisamment détaillée ou incomplète. Votre aide est la bienvenue !
Bibliographie
- (en) Cramer W, Kicklighter D, Bondeau A et al. (1999) Comparing global models of terrestrial net primary productivity (NPP): overview and key results. Global Change Biology, 5, 1–15.
 Portail de l’écologie
Portail de l’écologie  Portail de la biologie marine
Portail de la biologie marine  Portail du monde maritime
Portail du monde maritime