
{kind=link}
Bryophyte

Parmi les plantes actuelles, les bryophytes terrestres et les bryophytes aquatiques sont celles qui ont conservé le plus de caractères des premières plantes ayant colonisé la terre ferme. Les ancêtres de toutes les plantes terrestres, donc des Bryophytes, sont des algues vertes charophycées. Même si certaines structures anatomiques leur permettent de résister à la dessiccation, les Bryophytes sont encore extrêmement dépendantes de l'eau ou des milieux humides, ou en tout cas d'une hygrométrie minimale au moment de leur reproduction.
Pris au sens large, c'est-à-dire celui des classifications traditionnelles, le terme bryophyte s'applique aux trois embranchements de plantes terrestres qui ne possèdent pas de vrai système vasculaire. Au sens strict, l'embranchement actuel des Bryophyta ne concerne que les mousses et les sphaignes au sens botanique strict (à l'exception donc des Hepaticophyta et des Anthocerotophyta).
Dans une certaine mesure, en empêchant physiquement des graines d'accéder au sol ou en limitant l'accès de la lumière aux graines déjà présentes dans les sols (spectre du rouge notamment, qui déclenche la germination de certaines graines) et/ou par allélopathie, les tapis de mousses peuvent éliminer ou sélectionner certaines espèces[1] ; par exemple dans une pelouse calcaire néerlandaise (et de même en conditions contrôlées en serre), un tapis dense de mousses sur le sol réduit jusqu'à 30 % le nombre de plantules de certaines espèces phanérogames[1].
Cycle de vie des bryophytes




Comme pour les algues vertes et les plantes vertes, le cycle de vie présente une alternance de sporophytes (ou sporogone)et de gamétophytes, mais, comme pour toutes les Embryophytes, les anthérozoïdes (ou spermatozoïdes) sont protégés (et formés) par des anthéridies et les oosphères par des archégones (caractères partagés avec les plantes vasculaires).
La fécondation est aquatique (oogamie), les anthérozoïdes (ou spermatozoïdes) ciliés doivent trouver un moyen de rejoindre les oosphères. L'embryon qui en résulte est nourri et protégé par le gamétophyte (matrotrophie). Le sporophyte ne devient jamais indépendant de celui-ci. Le gamétophyte haploïde est donc le stade dominant sous lequel se rencontre la plante. Les spores sont entourées d'une paroi contenant de la sporopollénine qui les protège au cours de la dissémination. On parle d'un cycle génétique haplodiplophasique digénétique (présence de 2 générations séparées , le gamétophyte et le sporophyte) avec prédominance du gamétophyte sur le sporophyte (sporogone).
Organisation fonctionnelle
Les Bryophytes ne disposent pas de xylème ni de phloème (tissus conducteurs de l'eau et des nutriments). Ces deux tissus ne sont apparus que plus tard au cours de l'évolution, chez les plantes vasculaires. Certains bryophytes disposent cependant de tissus conducteurs spécialisés, mais ces derniers ne sont pas lignifiés : les « leptoïdes » et les « hydroïdes ».
Les pseudophylles n'ont pas l'histologie des vraies feuilles.
Il n'existe pas d'organes comparables aux racines chez les bryophytes mais des organes « rhizoïdes » (mono ou pluricellulaires) les fixent au substrat mais n'absorbent pas la solution du sol. Ces pseudos racines sont en fait d'anciens cormus (tige + feuille) qui en l'absence de photosynthèse se nécrose, se décolore et forment ainsi un humus primitif.
Les bryophytes ne pouvant absorber l'eau par leur rhizoÏde ont la faculté de permettre a l'eau de rentrer par toutes les parties de la plante. De plus en cas de forte sècheresse la plante peut entrer en état de vie ralentie (reviviscence) en attendant des conditions de développement mieux adaptées.
Comme les plantes vascularisées, les bryophytes peuvent profiter d'une symbiose avec des champignons.
Symbioses avec des champignons
On a encore peu de connaissances sur la diversité fongique associée aux bryophytes. Mais quelques études basées sur la biologie moléculaire ont montré qu'elle était importante.
Par exemple, en forêt boréale les bryophytes se montrent très résistantes au froid et capables de se nourrir sur des sols gelés tout ou partie de l'année. Les champignons les y aident. Il existe au nord du Canada « un ensemble écologiquement et phylogénétiquement diversifié de champignons associés aux parties vivantes de 3 bryophytes abondantes et largement distribuées (...),les Hylocomium splendens (Hedw.) Schimp. in B.S.G., Pleurozium schreberi (Brid.) Mitt. et Polytrichum commune Hedw »[2] ; Au moins 158 espèces de champignons leur sont associées dans la région étudiée. Ces champignons appartiennent surtout à l'embranchement des Ascomycota (62,8 % des cas), et à la subdivision des Basidiomycota (32 % des cas), parfois des Chytridiomycota (considérés comme les ancêtres de tous les autres champignons (3,9 % des cas) et rarement aux Glomeromycota (1,3 % seulement des cas, bien que ces champignons soient très fréquemment associés aux plantes supérieures)[2]. Les principaux ordres concernés étaient dans cette région les Hélotiales (18,6 %), les Agaricales (11,5 %), les Chaetothyriales (9,6 %) et les Tremellales[2]. L'association la plus souvent repérée était l'agaric Entoloma conferendum avec Lophodermium piceae (Fckl.) Hoehn. comme endophyte associé[2]. D'autres taxons avaient des affinités avec des Hélotiales du genre Hyphodiscus ou avec plusieurs basidiomycètes mycorhiziens[2]. La plupart (72,2 %) des taxons de champignons endophytes n'ont été trouvés qu'une seule fois dans cette étude[2]. De plus, très peu de champignons étaient associés avec l'ensemble des 3 bryophytes étudiés, ce qui indique une diversité d'associations très large et peut-être certaines spécificités[2], qui restent à explorer.
Habitats et répartition
Capables de tirer leurs nutriments de l'air et des aérosols, comme les lichens qu'ils accompagnent souvent, les bryophytes sont en tant que groupe taxonomique très ubiquistes. Grâce à de nombreuses adaptations, ils ont colonisé presque toute la planète, jusqu'aux zones subpolaires[3]. Ils sont toutefois absents (plus que les lichens) du milieu marin, et des milieux extrêmement arides. Grâce à des spores aéroportées, ils comptent parmi les premières espèces à pouvoir coloniser les nouveaux habitats (ex : terrils, îles volcaniques nouvelles...)
De nombreuses espèces vivent sur le sol, d'autres sont saxicoles (ou devenues muricoles) ou épiphytes, ou encore se développent sur les bois-morts tombés au sol. Dans les réserves naturelles et en forêt dans les îlots de sénescence, après une vingtaine d'années de non-gestion (« restauration passive »[4]), le taux de bois mort au sol ré-augmente au profit des bryophytes notamment[4] mais aussi de nombreux cortèges saproxylophages, chacun caractéristique de l'un des stades de décomposition du bois, et du type d'essences (bois dur, bois tendres, etc) ou de situation (à l'ombre, au soleil, dans l'eau...).
Nous pouvons également souligner un fait déroutant; au Canada, une chercheuse est parvenue à faire revivre en laboratoire des bryophytes prisonnières d'un glacier depuis pratiquement 400 ans. En exposant simplement ces plantes au soleil et à l'air, les bryophytes peuvent donc "ressusciter" après une très longue période d'emprisonnement dans la glace. Cette découverte permet de prouver l'étonnante résistance des bryophytes, alors que la communauté scientifique estimait qu'aucune plante ne pouvait survivre à une telle situation.
Cycle de vie
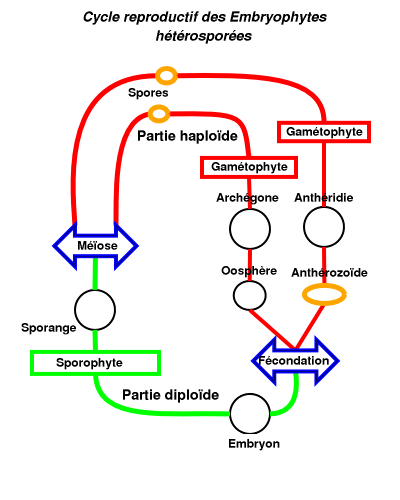
- La plus grande partie du cycle de vie s'effectue sous la forme de gamétophytes mâles ou femelles (en rouge). Dans certains cas, en particulier chez les anthocérotes, les gamétophytes sont bisexués et nourrissent alors de nombreux sporophytes.
- L'eau est indispensable au moment de la fécondation pour que les anthérozoïdes (ou spermatozoïdes) ciliés puissent atteindre les oosphères.
- Après la fécondation, l'embryon matotrophe est porté par le gamétophyte femelle et donne un sporophyte souvent formé d'une soie et d'un sporange (en vert).
- Le sporange libère des spores mâles et femelles (sauf dans le cas des gamétophytes bisexués) qui, chez les mousses essentiellement, se développent sous forme de protonéma avant de produire les gamétophytes.
- Les stades mobiles (en jaune) sont principalement les spores pour lesquelles les mousses ont développé des mécanismes assurant leur dispersion (capsule explosive des sphaignes, fentes de déhiscence des andréales, péristome des vraies mousses) et les anthérozoïdes (ou spermatozoïdes) avant la fécondation.
- Les bryophytes peuvent se multiplier de façon végétative par fragmentation. Certaines hépatiques ont développé des structures spéciales dispersant des «propagules». Des structures particulières existent aussi chez certaines mousses.
Bryophyte, bioindication, environnement et pollution...
Les bryophytes, et les mousses en particulier tirent l'essentiel de leur nourriture de l'air et des nutriments apportés par l'eau[5], par la pluie ou les dépôts de particules et aérosols. De nombreux bryophytes sont vulnérables à l'acidification de l'air, à sa pollution[6] et à l'eutrophisation des milieux[7].
Ils jouent à ce titre un rôle important dans l'épuration de l'air, mais aussi en matière d'accumulation de certains polluants (métaux lourds et radionucléides notamment). Ce sont aussi des organismes pionniers important dans les processus de résilience écologique.
Ils sont moins sensibles à la continuité temporelle de l'état forestier que les lichens[8]
Les espèces les plus sensibles aux polluants acides et aux pesticides véhiculés par l'air et les pesticides ont parfois disparu de tout ou partie de leur habitat naturel. Certaines espèces pourraient être considérées comme des bioindicateurs, en matière de qualité thermohygrométrique de l'air notamment. De par leur résistance naturelle à de nombreux polluants ils ne font pas de bons bioindicateurs de pollution, mais comme bioaccumulateurs, ils peuvent permettent de cartographier la pollution présente et d'un passé récent. On les a utilisé par exemple pour cartographier des pollutions par le plomb ou le cadmium.
Ils ont aussi été utilisés comme indicateurs de changement climatique récent[9].
Classification des bryophytes
- Hepaticophyta (hépatiques), l'embranchement aux caractères les plus primitifs (6000 espèces environs)
- Marchantiopsida (hépatiques à thalle complexe)
- Jungermanniopsida (hépatiques à thalle simple ou à feuille; environ 4000 espèces)
- Anthocerotophyta (anthocérotes) petit embranchement voisin des hépatiques (une centaine d'espèces)
- Bryophyta (mousses), près de 10000 espèces en 3 classes
- Sphagnopsida (sphaignes)
- Andreaeopsida (andréales)
- Bryopsida (vraies mousses)
État, pression, menaces sur les populations de bryophytes
Certaines espèces ont disparu de régions entières, ou doivent être considérée comme rares, devenues rares menacées ou en danger (une liste rouge de bryophytes a par exemple été publiée pour le Luxembourg[10]
Voir aussi
Bibliographie
- Bibliographie sur les Bryophytes du site Tela-Botanica, très complète.
- Frey, W., Frahm, J.P., Fischer, E., & Lobin, W. (2006), The Liverworts, Mosses and Ferns of Europe. Harley Books, Colchester
- Goffinet, B., Buck, W.R., & Shaw, A.J. (2008), Morphology, anatomy and classification of the Bryophyta. Pages 55–138 of: B., Goffinet, & A.J., Shaw (eds), Bryophyte Biology second edition. Cambridge University Press.
- Prune PELLET - Bryoflore des pierriers gréseux du massif du Suquet (Aigoual, Cévennes), Répartition, inventaire, caractérisation bryocénotiques et écologiques et évaluation de l'intérêt patrimonial, Rapport de stage Parc National des Cévennes, MNHN. Mis en ligne par Tela-Botanica.
- Vincent HUGONNOT, Mousses et hépatiques de Païolive (Ardèche et Gard, France), Jarnac, Société Botanique du Centre-Ouest, , 293 p. (ISSN 0759-934X)
Article connexe
- Bryologie
- Tortula
- Bryophyte aquatique
Liens externes
- Bryophytes, article en ligne de l'Encyclopædia Universalis
- Les bryophytes, présentation du Centre de ressources pédagogiques en Biologie Végétale de l'Université de Bourgogne
- Référence ITIS : — Hepaticophyta (fr) ( (en))
- Référence ITIS : — Anthocerotophyta (fr) ( (en))
- Référence ITIS : — Bryophyta (fr) ( (en))
- Tree of life
- Atlas de la Flore de France produit par la Fédération des conservatoires botaniques nationaux avec des données de répartition sur quelques dizaines de bryophytes de France.
Références
- 1 2 Tooren, B. V. (1990). Effects of a bryophyte layer on the emergence of seedlings of chalk grassland species. Acta Oecologica, 11(2), 155-163 (résumé)
- 1 2 3 4 5 6 7 Håvard Kauserud, Cecilie Mathiesen, Mikael Ohlson, High diversity of fungi associated with living parts of boreal forest bryophytes Botanique, 2008, 86(11): 1326-1333, 10.1139/B08-102 (Résumé)
- ↑ Longton, R.E. (1988), Biology of polar bryophytes and lichens. British Bryological Society Cambridge University Press..
- 1 2 Meyer, P., & Schmidt, M. 2010. Accumulation of dead wood in abandoned beech (Fagus sylvatica L.) forests in northwestern Germany. Forest Ecology and Management, 261, 343–352 (résumé).
- ↑ Kooijman AM, Bakker C. (1995), Species replacement in the bryophyte layer in mires: the role of water type, nutrient supply and interspecific interactions ; Journal of Ecology 83: 1–8. (résumé)
- ↑ Kooijman AM, Bakker C. (1994), The acidification capacity of wetland bryophytes as influenced by clean and polluted rain. Aquatic Botany 48: 133–144.
- ↑ Kooijman AM. 1993. Changes in the bryophyte layer of rich fens as controlled by acidification and eutrophication. PhD Thesis, Utrecht University, the Netherlands
- ↑ Fritz, Ö., Gustafsson, L., & Larsson, K. (2008), Does forest continuity matter in conservation? - A study of epiphytic lichens and bryophytes in beech forests of Southern Sweden. Biological Conservation, 141, 655–668. (résumé et graphiques)
- ↑ Frahm, J.P., & Klaus, D. (2001), Bryophytes as indicators of recent climate fluctuations in Central Europe. Lindbergia, 26(2), 97–104
- ↑ Werner J (1987) Liste rouge des bryophytes du Grand-Duché de Luxembourg. Musée d'histoire naturelle (PDF, 46 p).
 Portail de l’évolution
Portail de l’évolution  Portail de la botanique
Portail de la botanique