
{kind=link}

Deinonychus
?? propos de ce ??coles s??lection Wikipedia
SOS Enfants a essay?? de rendre le contenu plus accessible Wikipedia par cette s??lection des ??coles. Les enfants SOS est le plus grand don de charit?? du monde enfants orphelins et abandonn??s la chance de la vie familiale.
| Deinonychus Plage temporelle: Cr??tac?? inf??rieur, 115-108Ma | |
|---|---|
 | |
| Squelette mont??, Field Museum | |
| Classification scientifique | |
| Uni: | Animalia |
| Embranchement: | Chordata |
| Classe: | Reptilia |
| Ordre: | Saurischia |
| Famille: | ??? Dromaeosauridae |
| Clade: | ??? Eudromaeosauria |
| Sous-famille: | ??? Velociraptorinae |
| Genre: | ??? Deinonychus Ostrom 1969, |
| Esp??ce: | ??? D. antirrhopus |
| Nom binomial | |
| antirrhopus de Deinonychus Ostrom 1969, | |
| Synonymes | |
Velociraptor antirrhopus | |
Deinonychus (pron .: / r?? aɪ n ɒ n ɨ k ə s / DY- -ikəs NON; grec : δεινός, ??terrible?? et ὄνυξ, ὄνυχος g??nitif ??griffe??) est un genre de carnivores dromaeosaurid dinosaures . Il ya une esp??ce d??crite, Deinonychus antirrhopus. Ces dinosaures, qui pourrait atteindre jusqu'?? 3,4 m??tres (11 pieds) de long, a v??cu au d??but du Cr??tac?? P??riode, environ 115 ?? 108 il ya des millions d'ann??es (?? partir de la mi- Aptien au d??but Albian ??tapes). Les fossiles ont ??t?? r??cup??r??s de la ??tats am??ricains de Montana, Wyoming, et Oklahoma, dans les roches de la Formation et Cloverly Antlers Formation, si les dents qui peuvent appartenir ?? Deinonychus ont ??t?? trouv??s beaucoup plus ?? l'est dans Maryland.
Pal??ontologiste L'??tude de John Ostrom de Deinonychus ?? la fin des ann??es 1960 a r??volutionn?? la fa??on dont les scientifiques pensaient sur les dinosaures, conduisant ?? la " dinosaure renaissance "et allumer le d??bat ?? savoir si les dinosaures ??taient sang chaud ou sang froid. Avant cela, la conception populaire de dinosaures avait ??t?? l'un des laborieux, reptiles g??ants. Ostrom a not?? le petit corps, ??l??gant, position horizontale, ratites comme la colonne vert??brale, et surtout les griffes rapaces ??largis sur les pieds, qui sugg??raient, un pr??dateur agile active.
"Griffe terrible" se r??f??re ?? la inhabituellement ??lev??, talon en forme de croissant sur le deuxi??me orteil de chaque patte post??rieure. Le fossile YPM 5205 conserve une grande, fortement courb??e ungu??al. Dans la vie, archosaurs ont un ??tui corn?? sur cet os, qui se ??tend de la longueur. Ostrom regarda crocodiles et oiseaux griffes et reconstruit la griffe pour YPM 5205 que plus de 120 millim??tres (4,7 po) de long. Le nom de l'esp??ce antirrhopus signifie ??contrepoids??, qui renvoie ?? l'id??e de Ostrom sur la fonction de la queue. Comme dans d'autres dromaeosaurids, la queue ont vert??bres une s??rie de tendons ossifi??s et os allong?? super- proc??d??s. Ces caract??ristiques semblait faire la queue dans un contrepoids raide, mais un fossile de la tr??s ??troitement li??e mongoliensis de Velociraptor (IGM 100/986) a une queue squelette articul?? qui est courb?? lat??ralement dans une forme en S longtemps. Cela sugg??re que, dans la vie, la queue pourrait plier aux c??t??s avec un haut degr?? de flexibilit??.
Dans les deux formations Cloverly et Antlers, Deinonychus reste ont ??t?? trouv??s ??troitement associ??s ?? ceux de l'ornithopode Tenontosaurus. Dents d??couverts associ??s ?? des sp??cimens Tenontosaurus impliquent qu'ils ont ??t?? chass??s, ou au moins sur balay??s par Deinonychus.
Description

Bas?? sur les sp??cimens plus grand connues, Deinonychus pourrait atteindre au moins 3,4 m??tres (11 pi 2 po) de longueur, avec une longueur du cr??ne de 410 mm (16,1 po), une hauteur de hanche de 0,87 m??tres (2 pi 10 po) et un poids estim?? de 73 kilogrammes (160 kg). Le cr??ne a ??t?? ??quip?? de puissantes m??choires bord??es de dents incurv??es environ soixante-dix, lame-like. ??tudes du cr??ne ont beaucoup progress?? au cours des d??cennies. Ostrom reconstruit les partielles, cr??nes mal conserv??s qu'il avait comme triangulaire, large et assez semblable ?? Allosaurus . Autres documents de cr??ne Deinonychus et des esp??ces ??troitement li??es trouv??es avec une bonne conservation en trois dimensions montrent que le palais ??tait plus vo??t?? que Ostrom pensait, ce qui rend le museau beaucoup plus ??troite, tandis que le jugals ??vas??es largement, donnant une plus grande vision st??r??oscopique. Le cr??ne de Deinonychus ??tait diff??rente de celle de Velociraptor, cependant, en ce qu'il a un toit cr??nien plus robuste, comme celle de Dromaeosaurus, et ne avait pas les nasales d??prim??s de Velociraptor. Tant le cr??ne et la m??choire inf??rieure avaient fenestrae (Les ouvertures du cr??ne) qui a r??duit le poids du cr??ne. Dans Deinonychus, le fen??tre ant??orbitaire, une ouverture du cr??ne entre l'??il et la narine, a ??t?? particuli??rement importante.
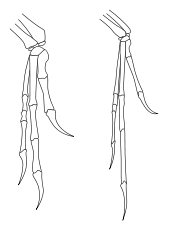
Deinonychus poss??dait de grandes ??mains?? ( manus) avec trois griffes sur chaque patte avant. Le premier chiffre est la plus courte et la deuxi??me a ??t?? plus longue. Chaque pied arri??re portait une griffe en forme de faucille sur le deuxi??me chiffre, qui a probablement ??t?? utilis?? pendant la pr??dation.
Aucun impressions de peau ont jamais ??t?? trouv??s en association avec des fossiles de Deinonychus. N??anmoins, la preuve sugg??re que le Dromaeosauridae, y compris Deinonychus, avait des plumes. Le genre Microraptor est ?? la fois phylog??n??tiquement plus g??ologiquement et plus primitive que Deinonychus, et au sein de la m??me famille. Plusieurs fossiles de Microraptor pr??server pennaceous, plumes ?? ailettes comme ceux des oiseaux modernes sur les bras, les jambes et la queue, avec plumes de couverture et de contour. Velociraptor est g??ologiquement plus jeune que Deinonychus, mais encore plus ??troitement li??s (au sein de la sous-famille Velociraptorinae, voir la classification , ci-dessous). Un sp??cimen de Velociraptor a ??t?? trouv?? avec les boutons d'oie sur le cubitus. Quill boutons sont o?? les ligaments folliculaires attach??s, et sont un indicateur direct de plumes d'aspect moderne.
Classification

Deinonychus antirrhopus est l'un des plus connus dromaeosaurid esp??ces, et est un proche parent de la petite Velociraptor , a trouv?? chez les jeunes, ?? la fin des formations rocheuses Cr??tac??-??ge en Asie centrale. Le clade qu'ils forment est appel?? Velociraptorinae. Sous le nom Velociraptorinae a ??t?? invent?? par Rinchen Barsbold en 1983 et ?? l'origine contenait le seul genre Velociraptor. Plus tard Phil Currie inclus la plupart des dromaeosaurids. Deux genres du Cr??tac?? sup??rieur, Tsaagan de Mongolie et d'Am??rique du Nord Saurornitholestes, peuvent ??galement ??tre proches parents, mais celui-ci est mal connue et difficile ?? classer. Velociraptor et ses alli??s sont consid??r??s comme utilisant leurs griffes plus que leurs cr??nes comme outils de tuer, plut??t que comme dromaeosaurids Dromaeosaurus, qui ont cr??nes trapu. En collaboration avec le troodontid??s, les dromaeosaurids forment la Deinonychosauria clade, qui est un taxon s??ur de aves . Phylog??n??tiquement, le Deinonychosauria repr??sentent le groupe de dinosaures non-aviaires les plus ??troitement li??es aux oiseaux.
D??couverte et nommage
Restes fossilis??s de Deinonychus ont ??t?? r??cup??r??s ?? partir de la Formation des Cloverly Montana et Wyoming et dans le plus ou moins contemporain Formation de Antlers Oklahoma, en Am??rique du Nord . La formation Cloverly a ??t?? dat?? de la fin Aptien jusqu'au d??but Albien du d??but du Cr??tac?? , environ 115 ?? 108 Ma. En outre, les dents trouv??es dans la Arundel Clay Faci??s (mi-Aptien), de la La formation sur le Potomac La plaine c??ti??re atlantique du Maryland peut ??tre attribu?? au genre.
Les premiers restes ont ??t?? d??couverts en 1931 dans le sud Montana pr??s de la ville de Billings. Le chef d'??quipe, pal??ontologue Barnum Brown, ??tait principalement pr??occup?? par l'excavation et la pr??paration des restes de la dinosaure ornithopode Tenontosaurus, mais dans son rapport sur le terrain du site de fouilles ?? la Mus??e am??ricain d'histoire naturelle, il a signal?? la d??couverte d'un petit dinosaure carnivore ?? proximit?? d'un squelette Tenontosaurus ??, mais enferm?? dans la chaux difficile ?? pr??parer." Il a appel?? de mani??re informelle l'animal "Daptosaurus agilis" et a fait des pr??paratifs pour d??crire et ayant le squelette, sp??cimen AMNH 3015, mettre sur l'??cran, mais jamais termin?? ce travail. Brown ramen?? de la Formation Cloverly le squelette d'un th??ropode plus petit avec des dents apparemment surdimensionn??es qu'il officieusement nomm?? "Megadontosaurus". John Ostrom, commentez ce mat??riau d??cennies plus tard, se rendit compte que les dents sont venus de Deinonychus, mais le squelette est venu d'un animal compl??tement diff??rent. Il a nomm?? ce squelette Microvenator.
Un peu plus de trente ans plus tard, en Ao??t 1964, le pal??ontologue John Ostrom a men?? une exp??dition du Mus??e Peabody de l'Universit?? de Yale qui a d??couvert mat??riau plus pr??s du squelette Bridger. Exp??ditions pendant les deux ??t??s suivants d??couvert plus de 1000 os, parmi lesquels au moins trois personnes. Depuis l'association entre les diff??rents os r??cup??r??s ??tait faible, ce qui rend le nombre exact d'animaux individuels repr??sent?? impossible de d??terminer correctement, le Nature de l'??chantillon (YPM 5205) de Deinonychus a ??t?? limit?? au pied gauche et du pied compl??te partielle droite qui a certainement appartenu ?? la m??me personne. Les ??chantillons restants ont ??t?? catalogu??s dans cinquante entr??es s??par??es au Yale Peabody Museum of Natural History.

??tude ult??rieure par Ostrom et Grant E. Meyer analys?? leur propre mat??riel ainsi que "Daptosaurus" de Brown en d??tail et a conclu qu'ils ??taient les m??mes esp??ces. Ostrom abord publi?? ses conclusions en F??vrier 1969, donnant tout le reste vis?? le nouveau nom de Deinonychus antirrhopus. Le nom sp??cifique "antirrhopus", ?? partir ἀντίρροπος grec, signifie ??contrepoids?? et fait r??f??rence ?? l'objet susceptible d'une queue raidie. En Juillet 1969, Ostrom a publi?? un tr??s vaste monographie sur Deinonychus.
Bien que d'une myriade d'os ??tait disponible en 1969, beaucoup de ceux importantes manquaient ou ??taient difficiles ?? interpr??ter. Il y avait quelques ??l??ments sus-orbitaires du cr??ne, pas de f??murs, pas sacrum, ne furcula ou du sternum, vert??bres manquant, et (Ostrom pensait) seule une infime fragment d'un coraco??de. La reconstruction du squelette de Ostrom de Deinonychus inclus un os tr??s inhabituel pelvienne - un pubis qui ??tait trap??zo??dale et plat, contrairement ?? celle des autres th??ropodes, mais qui ??tait la m??me longueur que l'ischion et qui a ??t?? trouv?? juste ?? c??t??.
D'autres r??sultats
En 1974, Ostrom a publi?? une monographie sur l'??paule de Deinonychus dans lequel il a r??alis?? que le pubis qu'il avait d??crit ??tait en fait un coraco??de - un ??l??ment de l'??paule. Dans la m??me ann??e, un autre sp??cimen de Deinonychus, MCZ 4371, a ??t?? excav??es dans le Montana par une exp??dition dirig??e par l'Universit?? de Harvard Farish Jenkins. Cette d??couverte a ajout?? plusieurs nouveaux ??l??ments: f??murs bien conserv??s, pubis, un sacrum, et de meilleures Ilia, ainsi que des ??l??ments du PSE et m??tatarse. Ostrom d??crit ce sp??cimen et r??vis?? sa restauration squelettique Deinonychus. Cette fois, il a montr?? la tr??s longue pubis, et Ostrom a commenc?? ?? soup??onner qu'ils peuvent ont m??me ??t?? un peu r??troversion comme ceux des oiseaux.
Un squelette de Deinonychus, y compris les os de l'original (et la plus compl??te) AMNH 3015 sp??cimen, peut ??tre vu sur l'??cran ?? l'American Museum of Natural History, avec un autre sp??cimen expos??s au Mus??e de zoologie compar??e ?? Universit?? De Harvard. Les sp??cimens American Museum et Harvard sont d'une localit?? diff??rente de celle des sp??cimens de Yale. M??me ces deux supports squelettiques sont des ??l??ments, y compris le Sterna, les c??tes du sternum, furcula et gastralia d??faut.
M??me apr??s tous les travaux d'Ostrom, plusieurs petits blocs de mat??riau de chaux enferm?? restaient pas pr??par??s ?? l'entreposage ?? l'American Museum. Ceux-ci ont consist?? principalement d'os isol??s et des fragments d'os, y compris la matrice d'origine, ou de la roche environnante, dans lequel les sp??cimens ont ??t?? initialement enterr??s. L'examen de ces blocs pr??par??s par Gerald Grellet-Tinner et Peter Makovicky en 2000 a r??v??l??, une caract??ristique int??ressante vis ?? vis. Plusieurs os longs, minces identifi??s sur les blocs que tendons ossifi??s (structures qui ont contribu?? ?? durcir la queue de Deinonychus) se est av??r?? repr??senter effectivement gastralia (Les nervures abdominaux). Plus important encore, un grand nombre de coquilles d'??ufs fossilis??s pr??c??demment inaper??ues ont ??t?? d??couverts dans la matrice rocheuse qui avait entour?? l'??chantillon Deinonychus originaux.
Dans un rapport ult??rieur, plus d??taill??es sur les coquilles d'??ufs, Grellet-Tinner et Makovicky conclu que la oeuf presque certainement appartenu ?? Deinonychus, repr??sentant le premier ??uf dromaeosaurid ??tre identifi??s. En outre, la surface externe de la coquille a ??t?? trouv?? une en contact ??troit avec la gastralia sugg??rant que Deinonychus pourrait avoir couv?? ses ??ufs. Cela implique que Deinonychus utilis?? le transfert de la chaleur du corps en tant que m??canisme pour l'incubation des ??ufs, et indique une endothermie similaire ?? oiseaux modernes. Une ??tude plus approfondie par Gregory Erickson et ses coll??gues estime que cet individu ??tait ??g?? de 13 ou 14 ans au moment du d??c??s et sa croissance a atteint un plateau. Contrairement ?? d'autres th??ropodes dans leur ??tude des sp??cimens trouv??s associ??s avec des ??ufs ou des nids, il avait fini sa croissance au moment de sa mort.
Implications
La description de Ostrom de Deinonychus en 1969 a ??t?? d??crit comme la plus importante d??couverte unique de pal??ontologie des dinosaures dans le milieu du 20e si??cle. La d??couverte de ce pr??dateur agile clairement actif a fait beaucoup pour changer la conception scientifique (et populaire) des dinosaures et a ouvert la porte ?? la sp??culation que les dinosaures aient ??t?? sang chaud. Ce d??veloppement a ??t?? appel??e dinosaure renaissance. Plusieurs ann??es plus tard, Ostrom a not?? des similitudes entre les pattes ant??rieures de Deinonychus et que des oiseaux, une observation qui l'a amen?? ?? relancer l'hypoth??se que les oiseaux descendent des dinosaures. Quarante ans plus tard, cette id??e est presque universellement accept??e.
En raison de son anatomie tr??s ressemblant ?? des oiseaux et relation ??troite avec d'autres dromaeosaurids, les pal??ontologues ont ??mis l'hypoth??se que Deinonychus ??tait probablement recouvert de plumes. Preuve fossile claire de plumes aviaire modernes de style existe depuis plusieurs dromaeosaurids connexes, y compris Velociraptor et Microraptor, si aucune preuve directe ne est encore connu pour Deinonychus lui-m??me. Lorsque la r??alisation d'??tudes de domaines tels que l'amplitude du mouvement dans les pattes avant, les pal??ontologues comme Phil Senter ont pris la pr??sence probable de plumes des ailes (que pr??sente dans tous les dromaeosaurs connus avec des impressions de peau) en consid??ration.
Pal??obiologie et pal??o??cologie
Les donn??es g??ologiques indiquent que Deinonychus habit?? un des plaines inondables ou swamplike habitat. Le pal??oenvironnement la fois de la formation sup??rieure et la Cloverly Antlers Formation, dans lequel reste de Deinonychus ont ??t?? trouv??s, compos??e de for??ts tropicales ou sub-tropicales, deltas et des lagunes, un peu comme aujourd'hui Louisiane. Autres animaux Deinonychus partag?? son monde avec inclure des dinosaures herbivores comme le blind?? Sauropelta et les ornithopodes Zephyrosaurus et Tenontosaurus. Dans l'Oklahoma, l'??cosyst??me de Deinonychus ??galement inclus le grand th??ropode Acrocanthosaurus, l'??norme sauropode Sauroposeidon , le crocodilien Goniopholis, et la gar Lepisosteus. Si les dents trouv??es dans le Maryland sont celles de Deinonychus, puis ses voisins seraient comprennent Astrodon et un nodosaur (appel??e Priconodon) ne est connu que de dents. La partie m??diane de la Formation Cloverly varie dans l'??ge de 115 ?? 10 Ma pr??s de la base ?? 108,5 ?? 0,2 Ma pr??s du sommet. Dans une ??tude de 2001 men??e par Bruce Rothschild et d'autres pal??ontologues, 43 os de la main et 52 os du pied vis??es ?? Deinonychus ont ??t?? examin??s pour des signes de fracture de stress; aucune n'a ??t?? trouv??e.
Le comportement pr??dateur

Deinonychus dents trouv??s en association avec des fossiles de la Tenontosaurus de dinosaures ornithopodes sont assez fr??quents dans la formation Cloverly. Deux carri??res ont ??t?? d??couverts qui pr??servent fossiles assez complets Deinonychus pr??s fossiles Tenontosaurus. La premi??re, la carri??re de Yale dans le Cloverly du Montana, comprend de nombreuses dents, quatre adultes et un mineur Deinonychus Deinonychus. L'association de ce nombre de Deinonychus squelettes dans une seule carri??re sugg??re que Deinonychus ont peut-??tre nourri de cet animal, et peut-??tre chass?? il. Ostrom et Maxwell ont m??me utilis?? cette information pour sp??culer que Deinonychus pourrait avoir v??cu et chass?? en meute. La deuxi??me tels carri??re est de la Formation Antlers de l'Oklahoma. Le site contient six squelettes partiels de Tenontosaurus de diff??rentes tailles, avec une squelette partiel et de nombreuses dents de Deinonychus. Un hum??rus tenontosaur porte m??me ce qui pourrait ??tre des marques de dents de Deinonychus. Brinkman et al. (1998) soulignent que Deinonychus avait une masse de 70 ?? 100 kg d'adultes, alors que tenontosaurs adultes ??taient 1-4 tonnes. Un Deinonychus solitaire ne pouvait pas tuer un tenontosaur adulte, ce qui sugg??re que la chasse bloc est possible.
Une ??tude de 2007 par Roach et Brinkman a remis en question le comportement coop??ratif de chasse de paquet de Deinonychus, sur la base de ce qui est connu de la chasse moderne et le carnivore taphonomie des sites de tenontosaur. Moderne archosaurs ( oiseaux et crocodiles ) et dragons de Komodo affichent peu de chasse coop??rative; ?? la place, ils sont g??n??ralement soit des chasseurs solitaires, ou sont attir??s par les carcasses pr??c??demment tu??s, o?? une grande partie de conflit entre individus de la m??me esp??ce. Par exemple, dans des situations o?? des groupes de dragons de Komodo mangent ensemble, les plus grands individus mangent en premier et se attaquer petits Komodos qui tentent de nourrir; si le petit animal est tu??, il est cannibalis??. Lorsque cette information est appliqu??e aux sites tenontosaur, il semble que ce qui est trouv?? est compatible avec Deinonychus ayant une strat??gie d'alimentation komodo- ou crocodile-comme. Restes squelettiques Deinonychus trouv??s sur ces sites sont de jeunes adultes, avec des pi??ces manquantes coh??rente d'avoir ??t?? mang?? par d'autres Deinonychus. D'autre part, un document r??dig?? par Li et al. D??crit sites de piste avec un espacement de pied similaire et voies de roulement parall??les, ce qui implique un comportement gr??gaire d'emballage au lieu du comportement alimentaire non coordonn??e.
En 2011, Denver Fowler et ses coll??gues ont propos?? une nouvelle m??thode par laquelle Deinonychus et d'autres peuvent dromaeosaurs ont captur?? et retenu proies. Ce mod??le, connu sous le nom "rapace proie retenue" (RPR) mod??le de pr??dation, propose que Deinonychus tu?? sa proie d'une mani??re tr??s similaire ?? existante accipitrid oiseaux de proie : par sauter sur sa proie, l'??pinglant sous son poids de corps, et saisissant fermement avec les grandes griffes, en forme de faucille. Comme accipitrids, le dromaeosaur serait alors commencer ?? se nourrir de l'animal en vie, jusqu'?? ce qu'il est finalement d??c??d?? de la perte de sang et la d??faillance d'organes. Cette proposition est bas??e principalement sur des comparaisons entre la morphologie et les proportions des pieds et les jambes de dromaeosaurs ?? plusieurs groupes d'oiseaux de proie existantes ayant des comportements pr??dateurs connus. Fowler a constat?? que les pieds et les jambes de dromaeosaurs ressemblent le plus ?? ceux de aigles et faucons , en particulier en termes d'avoir une seconde griffe ??largie et une gamme similaire de saisir le mouvement. Cependant, la courte m??tatarse et la force du pied auraient ??t?? plus semblable ?? celle des hiboux . La m??thode de RPR de pr??dation serait compatible avec d'autres aspects de l'anatomie Deinonychus s ??, comme la m??choire et le bras inhabituelle morphologie. Les bras ??taient probablement couvertes de longues plumes, et peuvent avoir ??t?? utilis??s comme stabilisateurs de battement pour l'??quilibre tout au sommet d'un animal de proie du mal, avec la queue contrepoids raide. Les m??choires, la pens??e d'avoir eu une force de morsure relativement faible, auraient ??t?? utiles pour manger la proie vivante, mais pas aussi utile pour rapide, l'envoi en force de la proie (puisque les animaux modernes qui le font ont sp??cialisations uniques qui dromaeosaurs manquaient). Ces adaptations pr??dateurs qui travaillent ensemble peuvent ??galement avoir des implications pour le origine de claquer au paravians.
force de morsure

Estimations de la population de morsure pour Deinonychus ont ??t?? produites la premi??re fois en 2005, sur la base de la m??choire reconstruit musculature. Cette ??tude a conclu que Deinonychus avait probablement une force de morsure maximale que seulement 15% de la modernit?? Alligator am??ricain. Une ??tude de 2010 par Paul Gignac et ses coll??gues a tent?? d'estimer la force de morsure fond??e directement sur les marques nouvellement d??couverts Deinonychus dent de ponction dans les os d'un Tenontosaurus. Ces marques de piq??res provenaient d'une grande personne, et fourni la premi??re preuve que les grandes Deinonychus pourraient mordre ?? travers l'os. En utilisant les marques de dents, l'??quipe de Gignac ont pu d??terminer que la force de morsure de Deinonychus ??tait significativement plus ??lev??e que les ??tudes ant??rieures avaient estim?? par des ??tudes biom??caniques seul. Ils ont trouv?? la force de morsure de Deinonychus se situer entre 4100 et 8200 newtons, plus que de vivre mammif??res carnivores dont l' hy??ne , et ??quivalent ?? un alligator de taille similaire.
Gignac et ses coll??gues ont ??galement not??, cependant, que les marques perforation d'os de Deinonychus sont relativement rares, et contrairement aux grandes th??ropodes avec de nombreuses marques de ponction connus comme Tyrannosaurus , Deinonychus ne ont probablement pas mordent souvent par ou mangent os. Au lieu de cela, ils ont probablement utilis?? leur force de morsure ??lev?? dans la d??fense ou la capture des proies, plut??t que de nourrir.
la fonction du membre

En d??pit d'??tre la caract??ristique la plus distinctive de Deinonychus, la forme et la courbure de la griffe faucille varie entre ??chantillons. Le sp??cimen type d??crit par Ostrom en 1969 a une pince faucille fortement courb??e, tout un sp??cimen nouveaux d??crits en 1976 avaient une griffe avec une courbure beaucoup plus faible, plus semblable au profil avec les griffes ??normales?? sur les orteils restants. Ostrom a sugg??r?? que cette diff??rence dans la taille et la forme des griffes de faucille pourrait ??tre due ?? la variation individuelle, sexuelle ou li??e ?? l'??ge, mais il a admis qu'il ne pouvait pas en ??tre s??r.
Il existe des preuves anatomiques et que cette voie de circulation Talon a eu lieu au large de la terre tandis que le dinosaure marchait sur les troisi??me et quatri??me orteils.
Ostrom a sugg??r?? que Deinonychus pourrait botter avec la griffe faucille pour couper et r??duire ?? sa proie. Certains chercheurs ont m??me sugg??r?? que le talon a ??t?? utilis?? pour ??ventrer grande dinosaures c??ratopsiens. D'autres ??tudes ont sugg??r?? que les griffes de faucille ne ont pas ??t?? utilis??s pour r??duire mais plut??t pour offrir de petits coups ?? la victime. Manning et al. (2005) a effectu?? des essais sur une r??plique robotique qui correspondent pr??cis??ment l'anatomie de Deinonychus et Velociraptor , et utilis?? des v??rins hydrauliques pour faire le robot frapper une carcasse de porc. Dans ces essais, les serres ne ont fait que peu profondes perforations et ne pouvaient pas couper ou r??duire. Les auteurs sugg??rent que les serres auraient ??t?? plus efficace dans l'escalade que dans le traitement des coups de tuer.
Ostrom compar?? Deinonychus ?? l' autruche et le casoar . Il a not?? que les esp??ces d'oiseaux peuvent infliger de graves blessures ?? la grande griffe sur le deuxi??me orteil. Le casoar a des griffes jusqu'?? 125 millim??tres (4,9 po) de long. Ostrom cit?? Gilliard (1958) ?? dire qu'ils peuvent couper un bras ou ??ventrer un homme. Kofron (1999 et 2003) a ??tudi?? 241 attaques de casoar document??es et a constat?? que l'un deux chiens humaine et avaient ??t?? tu??s, mais aucune preuve que les casoars peuvent ??ventrer ou d??membrer autres animaux. Casoars utilisent leurs griffes pour se d??fendre, pour attaquer les animaux en danger, et affiche agonistes tels que la menace affichage cordes. Le seriema a ??galement une seconde griffe d'orteil ??largie, et l'utilise pour d??chirer petites proies pour les avaler. En 2011, une ??tude a sugg??r?? que la griffe faucille aurait probablement ??t?? utilis?? pour cerner proie tout en mordant, plut??t que comme une arme de d??broussaillage.
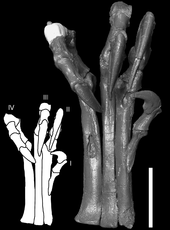
Des ??tudes biom??caniques par Ken Carpenter en 2002 a confirm?? que la fonction la plus probable des membres ant??rieurs ?? la pr??dation saisissait, comme leurs grands efforts auraient permis plus atteindre que pour la plupart des autres th??ropodes. Les rondeurs et allong?? coraco??de, indiquant muscles puissants dans les pattes avant, encore renforc?? cette interpr??tation. ??tudes biom??caniques de Carpenter partir de la moelle moulages ont ??galement montr?? que Deinonychus pouvait pas plier ses bras contre son corps comme un oiseau (??de pliage aviaire"), contrairement ?? ce qui a ??t?? d??duite des descriptions ant??rieures 1985 par Jacques Gauthier et Gregory S. Paul en 1988.
Des ??tudes men??es par Phil Senter en 2006 a indiqu?? que les membres ant??rieurs Deinonychus pourraient ??tre utilis??s non seulement pour saisir, mais aussi pour serrant les objets vers la poitrine. Si Deinonychus avait des doigts et des ailes de plumes, des plumes auraient limit?? l'amplitude de mouvement des membres ant??rieurs ?? un certain degr??. Par exemple, lorsque Deinonychus a ??tendu son bras vers l'avant, la ??palme?? de la main tourn?? automatiquement vers une position tourn??e vers le haut. Cela aurait provoqu?? une aile pour bloquer l'autre si les deux membres ant??rieurs ont ??t?? ??tendues en m??me temps, ce qui conduit ?? conclure que Senter objets embrayage ?? la poitrine ne aurait ??t?? accompli avec un bras ?? la fois. La fonction des doigts aurait ??galement ??t?? limit??e par des plumes; par exemple, seulement le troisi??me doigt de la main aurait pu ??tre employ??e dans des activit??s telles que le sondage crevasses pour les petites proies, et seulement dans une position perpendiculaire ?? l'aile principale. Alan Gishlick, dans une ??tude de 2001 de Deinonychus la m??canique des membres ant??rieurs, a constat?? que m??me si de grandes plumes des ailes ??taient pr??sents, la capacit?? de pr??hension de la main ne aurait pas ??t?? consid??rablement entrav??; plut??t, saisie aurait ??t?? accompli perpendiculaire ?? l'aile, et objets susceptibles aurait ??t?? tenue par les deux mains simultan??ment dans un mode "c??lin", les r??sultats qui ont ??t?? pris en charge par les ??tudes des membres ant??rieurs plus tard par Carpenter et Senter.
Parsons et Parsons ont montr?? que les mineurs et sous-adultes sp??cimens de Deinonychus affichent quelques diff??rences morphologiques avec les adultes. Par exemple, les bras des sp??cimens plus jeunes ??taient proportionnellement plus longues que celles des adultes, une indication possible de la diff??rence de comportement entre les jeunes et les adultes. Un autre exemple de ce qui pourrait ??tre la fonction des griffes de p??dale. Parsons et Parsons ont sugg??r?? que la courbure de la griffe (qui Ostrom [1976] avait d??j?? montr?? ??tait diff??rente entre les sp??cimens) ??tait peut-??tre plus important pour les mineurs Deinonychus, car cela pourrait aider ?? grimper dans les arbres, et que les griffes est devenu plus droit que l'animal est devenu plus et a commenc?? ?? vivre uniquement sur le terrain. Cette d??cision ??tait fond??e sur l'hypoth??se que certains petits dromaeosaurids utilis??s leurs griffes ?? p??dales pour l'escalade.
Vitesse

Dromaeosaurids, en particulier Deinonychus, sont souvent d??peints comme des animaux exceptionnellement ?? fonctionnement rapide dans les m??dias populaires, et Ostrom se sp??cul?? que Deinonychus ??tait agiles dans sa description originale. Cependant, lors de la premi??re d??crite, une jambe compl??te de Deinonychus ne avait pas ??t?? trouv??, et la sp??culation sur les Ostrom la longueur de la f??mur (os de la jambe du haut) se est av??r?? plus tard avoir ??t?? surestim??. Dans une ??tude ult??rieure, Ostrom a not?? que le rapport du f??mur ?? la tibia (inf??rieur de l'os de la jambe) ne est pas aussi important pour d??terminer la vitesse par rapport ?? la longueur du pied et de la jambe. Chez les oiseaux agiles modernes, comme l' autruche , le ratio pied-tibia est 0,95. Dans dinosaures particuli??rement ?? fonctionnement rapide, comme Struthiomimus , le ratio est de 0,68, mais dans Deinonychus le rapport est 0,48. Ostrom a d??clar?? que la "seule conclusion raisonnable??, ce est que Deinonychus ne ??tait pas particuli??rement rapide par rapport ?? d'autres dinosaures, et certainement pas aussi vite que les oiseaux de voler modernes.
Le faible ratio pied de la jambe dans Deinonychus est d?? en partie ?? une inhabituellement court m??tatarse (os du pied sup??rieure). Le rapport est en fait plus grande dans les petits individus que dans les grandes. Ostrom a sugg??r?? que le m??tatarse court peut ??tre li??e ?? la fonction de la griffe de faucille, et utilis?? du fait qu'il semble se raccourcir comme individus ??g??s de support pour ce produit. Il a interpr??t?? toutes ces caract??ristiques - le deuxi??me orteil court avec la griffe ??largie, ?? court m??tatarse, etc. - que le soutien ?? l'utilisation de la jambe arri??re comme une arme offensive, o?? la griffe faucille frapperait bas et en arri??re, et la jambe recula et bas dans le m??me temps, le d??broussaillage et ?? la d??chirure ?? la proie. Ostrom a sugg??r?? que le court m??tatarse r??duit le stress global sur les os de la jambe lors d'une telle attaque, et a interpr??t?? la disposition inhabituelle des attaches musculaires dans la jambe Deinonychus comme support pour son id??e que un ensemble diff??rent de muscles a ??t?? utilis?? dans la course pr??datrice que dans la marche ou la course. Par cons??quent, Ostrom a conclu que les jambes de Deinonychus repr??sentaient un ??quilibre entre les adaptations de fonctionnement n??cessaires ?? un pr??dateur agile, et les caract??ristiques de r??duction du stress pour compenser son arme de pied unique.
Dans son ??tude de 1981 des empreintes de dinosaures canadiens, Richard Kool produit des estimations de vitesse de marche rugueuses sur la base de plusieurs voies de roulement faites par diff??rentes esp??ces dans le Formation Gething du Colombie Britannique. Kool estim?? une de ces voies de roulement, repr??sentant le ichnoesp??ces Irenichnites gracilis (qui peut avoir ??t?? faite par Deinonychus), d'avoir une vitesse de marche de 10,1 kilom??tres par heure (six milles par heure).
??ufs

L'identification, en 2000, d'un Deinonychus oeuf probable associ?? ?? l'un des sp??cimens originaux ont permis de comparer avec d'autres dinosaures th??ropodes en termes de structure de l'??uf, la nidification et la reproduction. Dans leur examen de l'??chantillon 2006, Grellet-Tinner et Makovicky examin?? la possibilit?? que le dromaeosaurid avait ??t?? l'alimentation sur l'oeuf, ou que les fragments d'??ufs avaient ??t?? associ??e avec le squelette Deinonychus par hasard. Ils ont rejet?? l'id??e que l'??uf avait ??t?? un repas pour le th??ropode, notant que les fragments ont ??t?? pris en sandwich entre les c??tes du ventre et des membres ant??rieurs des os, ce qui rend impossible qu'ils repr??sentaient contenu de l'estomac de l'animal. En outre, la mani??re dont l'??uf avait ??t?? ??cras??e et fragment??e a indiqu?? qu'il avait ??t?? intacte au moment de l'enterrement, et a ??t?? bris??e par le processus de fossilisation. L'id??e que l'??uf a ??t?? associ??e de fa??on al??atoire avec le dinosaure a ??galement ??t?? jug??e peu probable; les os entourant l'ovule ne avaient pas ??t?? diffus??e ou d??sarticul??, mais il est rest?? assez intacte par rapport ?? leurs positions dans la vie, ce qui indique que la zone autour et y compris l'??uf n'a pas ??t?? perturb?? pendant la conservation. Le fait que ces os ??taient les c??tes du ventre ( gastralia), qui sont tr??s rarement trouv??s articul??, soutenu cette interpr??tation. Toutes les preuves, selon Grellet-Tinner et Makovicky, indique que l'oeuf ??tait intacte sous le corps de la Deinonychus quand il a ??t?? enterr??. Il est possible que cela repr??sente comportement de couvaison ou imbrication dans Deinonychus similaire ?? celle observ??e dans le apparent?? troodontid??s et oviraptorid??s, ou que l'oeuf est en fait ?? l'int??rieur du oviducte lorsque l'animal est mort.
L'examen de la microstructure de l'??uf Deinonychus confirme qu'il appartenait ?? un th??ropode, car il partage des caract??ristiques avec d'autres oeufs de th??ropodes connue et montre dissemblances avec ornithischian et oeufs sauropodes. Par rapport aux autres th??ropodes maniraptoran, l'??uf de Deinonychus est plus semblable ?? ceux de oviraptorid??s qu'?? ceux de troodontid??s, en d??pit des ??tudes qui montrent que ces derniers sont plus ??troitement li??e ?? dromaeosaurids comme Deinonychus. Bien que l'oeuf a ??t?? trop mal broy?? pour d??terminer avec pr??cision la taille et le Grellet-Tinner et des estimations Makovicky un diam??tre d'environ 7 cm (2,7 in) sur la base de la largeur du canal pelvien ?? travers laquelle l'??uf doit avoir r??ussi. Cette taille est similaire au diam??tre de 7,2 cm de la plus grande Citipati (un oviraptorid??s) oeufs; Citipati et Deinonychus a ??galement partag?? la m??me taille globale de corps, en soutenant cette estimation. En outre, les ??paisseurs des Citipati et Deinonychus coquilles d'??ufs sont presque identiques, et puisque l'??paisseur de la coque est en corr??lation avec le volume de l'??uf, ce soutient en outre l'id??e que les oeufs de ces deux animaux ??taient de la m??me taille.
Pathologie
La seconde phalange de l'orteil dans la deuxi??me sp??cimen YPM 5205 a une fracture gu??rit.