{kind=link}

Extinción masiva del Cretácico-Terciario
Acerca de este escuelas selección Wikipedia
SOS Children ha intentado que el contenido de Wikipedia más accesible por esta selección escuelas. Con Infantil SOS se puede elegir a apadrinar a los niños en más de cien países



El evento de extinción del Cretácico-Paleógeno ocurrió hace aproximadamente 65 hasta 66 millones años ( Ma) al final de la Edad Maastrichtiano del Cretácico período. Fue una intensa, global extinción en masa, se calcula que el 75% de los animales y las plantas, sobre todo no aviares dinosaurios , que tiene lugar en un geológicamente corto período de tiempo. A menudo se llama la extinción K-Pg, siendo K la abreviatura del término alemán para el Cretácico, Kreide, y Pg es la abreviatura para el Paleógeno. Es quizás el más conocido como el Cretácico-Terciario (KT), pero el término Terciario está desalentado como una unidad formal de la Comisión Internacional de Estratigrafía.
La extinción K-Pg puso fin a la Era Mesozoica y comenzó la Era Cenozoica, y se caracteriza por una fina capa de sedimentos, la K-Pg límite (o límite KT) de arcilla, que se encuentra en todo el mundo en rocas marinas y terrestres. La arcilla del límite muestra altos niveles de metal iridio , que es raro en la corteza de la Tierra pero abundante en los asteroides. Esto condujo a la hipótesis de que la extinción como resultado del impacto de una asteroide gigante o un cometa que causó perturbaciones catastróficas para el medio ambiente, incluyendo el cierre temporal de la fotosíntesis de las plantas de la tierra y el plancton. La identificación de la 180 kilometros (110 millas) de ancho Cráter Chicxulub en México aportan pruebas concluyentes de que la arcilla del límite K-Pg representa escombros de un impacto de asteroide. El hecho de que las extinciones se produjeron al mismo tiempo que el impacto proporciona una fuerte evidencia de que la extinción K-Pg fue causada por el asteroide. Sin embargo, algunos científicos continúan argumentando que otros factores, como las erupciones volcánicas, el cambio climático, y / o el cambio del nivel del mar, causados o por lo menos han contribuido a la extinción.
Una amplia gama de especies perecieron en la extinción K-Pg. Las víctimas más conocidas son los dinosaurios no aviares. Sin embargo, la extinción también se estrelló contra otros organismos terrestres, incluyendo mamíferos, pterosaurios, aves, lagartos, insectos y plantas. En los océanos, la extinción K-Pg devastó los lagartos marinos gigantes ( Mosasauridae), plesiosaurios, peces, tiburones, moluscos (especialmente los ammonites ) y muchas especies de plancton. Se estima que el 75% o más de todas las especies de la Tierra desapareció. Sin embargo, la devastación causada por la extinción también brindó oportunidades evolutivas. A raíz de la extinción, muchos grupos se sometieron a radiaciones adaptativas notables. Mamíferos, en particular, se diversificaron en el Paleógeno, produciendo nuevas formas tales como caballos, ballenas, murciélagos y primates. Aves, peces y lagartos quizás también radiadas.
Patrones de extinción
{kind=link}
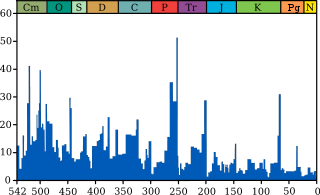
El evento de extinción K-Pg fue grave, global, rápida y selectiva. En cuanto a la gravedad, el evento eliminado un gran número de especies. Sobre la base de fósiles marinos, se estima que el 75% o más de todas las especies fueron exterminados por la extinción K-Pg. Esta es una estimación aproximada. Es difícil estimar la diversidad de los ecosistemas modernos, por no hablar de los fósiles, y los datos se deriva principalmente de invertebrados marinos. Los organismos terrestres, especialmente insectos, representan gran parte de la diversidad, pero tener un registro más pobre. A pesar de esto, los altos niveles de extinción observados en los fósiles terrestres y marinas indican que la extinción K-Pg es el más grave extinción en los últimos 250 millones de años.
La extinción K-Pg fue un evento global. El evento parece haber golpeado a todos los continentes a la vez. Los dinosaurios, por ejemplo, se conocen desde el Maastrichtiano de América del Norte, Europa, Asia, África, América del Sur y la Antártida, pero se desconoce desde el Cenozoico en cualquier parte del mundo. Del mismo modo, fósiles muestran polen devastación de las comunidades vegetales en áreas tan lejanas como Nuevo México, Alaska, China y Nueva Zelanda. El evento también afectó a todos los mares y océanos. Grupos generalizados como los mosasaurios y ammonites desaparecieron alrededor del mundo. Por otra parte, las extinciones se produjeron al mismo tiempo en la tierra y en el mar.
El registro fósil muestra que el ritmo de la extinción K-Pg era extremadamente rápido, que se producen en una escala de miles de años o menos. En algunos casos, es posible estudiar los fósiles en una escala muy fina - centímetro por centímetro - a través de las rocas K-Pg. Los ejemplos incluyen microfósiles marinos, como nanoplancton y foraminíferos calcáreos, y el polen de las plantas terrestres. Aquí, los fósiles muestran que el ecosistema se mantuvo relativamente estable hasta el límite K-Pg, momento en el que muchas especies desaparecen repentinamente. Para grupos con un registro fósil más pobres, como los dinosaurios, fósiles es poco probable que preservar justo por debajo del límite K-Pg. Por ejemplo, sólo unas pocas docenas de esqueletos Tyrannosaurus son conocidos, por lo que las probabilidades de encontrar uno a pocos centímetros por debajo de la frontera son bajos. Este efecto, denominado Efecto Signor-Lipps, hace que muchas especies parecen desvanecerse antes del límite K-Pg, creando la ilusión de la extinción gradual. Sin embargo, la mejora de muestreo indica que los grupos pensaban que someterse a un lento declive, como los dinosaurios, en realidad desaparecer repentinamente cerca del límite K-Pg. Volver a trabajar - cuando los fósiles se erosionan de rocas más antiguas y se depositan en rocas más jóvenes - también puede hacer que la extinción parece gradual. Por ejemplo, en los Bug Creek hormigueros camas en Montana, fósiles de dinosaurios ocurren junto mamíferos del Paleoceno temprano, lo que crea la ilusión de que los dinosaurios se redujeron como mamíferos radiadas. Fósiles reelaborados se reconocen porque tienden a ser poco frecuentes y, a menudo son dañados por la reelaboración.
Los patrones son fundamentales para la comprensión de la causa de las extinciones. El hecho de que la extinción es grave, global y rápida indica sugiere que las extinciones son el resultado de un disturbance- ambiental grave, global y rápida una catástrofe ambiental. Pero en los años 1970 y 1980, esto llevó a los científicos a considerar seriamente los mecanismos catastróficos como las supernovas, las erupciones volcánicas y los asteroides, y provocó un nuevo interés en catastrofismo en geología y paleontología.
La extinción también fue altamente selectiva. Algunos grupos eran relativamente poco afectada, otros fueron devastadas, y algunos fueron eliminados por completo. Muchas especies de cocodrilo, tortuga, y salamandra sobrevivieron, por ejemplo. Mamíferos, aves y lagartos sufrieron altas tasas de extinción. Dinosaurios no aviares y pterosaurios fueron exterminados por completo.
A pesar de que el evento límite fue grave, hay una variabilidad significativa en la tasa de extinción entre y dentro de diferentes clados. Las especies que dependen de la fotosíntesis se redujeron o se extinguieron como partículas atmosféricas bloquean la luz solar y reducen la energía solar que alcanza la superficie de la Tierra. Esta extinción de la planta provocó una importante reorganización de los grupos de plantas dominantes. Fotosíntesis organismos, incluyendo fitoplancton y plantas de la tierra, forman la base de la cadena alimentaria en el Cretácico tardío como lo hacen hoy. La evidencia sugiere que herbívoros animales se extinguieron cuando las plantas que dependían para la comida comenzó a escasear. En consecuencia, la parte superior depredadores como el Tyrannosaurus rex también perecieron.
Cocolitofóridos y moluscos (incluidos los ammonites , rudistas, de agua dulce caracoles y mejillones), y aquellos organismos cuyas cadena alimentaria incluyó estos constructores de cáscaras, se extinguió o sufrió graves pérdidas. Por ejemplo, se piensa que amonitas eran el principal alimento de mosasaurios, un grupo de marinos gigantescos reptiles que se extinguieron en el límite.
Los omnívoros, insectívoros y carroñeros sobrevivieron a la extinción, tal vez debido a la mayor disponibilidad de sus fuentes de alimento. Al final del Cretácico parece haber habido puramente herbívoros o carnívoros mamíferos. Los mamíferos y aves que sobrevivieron a la extinción alimentados con insectos , gusanos y caracoles, que a su vez se alimentan de plantas muertas y materia animal. Los científicos suponen que estos organismos sobrevivieron al colapso de las cadenas de comida basados en plantas porque se alimentaban de detritus (no vivos material orgánico).
En corriente comunidades pocos grupos de animales se extinguieron porque las comunidades corriente dependen menos directamente sobre los alimentos de las plantas vivas y más de detritus que se lava en el de la tierra, amortiguación de la extinción. Patrones similares, pero más complejas se han encontrado en los océanos. La extinción fue más grave entre los animales que viven en la columna de agua que entre los animales que viven sobre o en el fondo del mar. Los animales en la columna de agua dependen casi por completo de producción primaria de fitoplancton vida mientras que los animales que viven sobre o en el feed suelo marino de detritus o puede cambiar a la alimentación detritus.
Los mayores sobrevivientes de aspiración de aire del evento, crocodiliformes y champsosaurs, eran semi-acuático y tenía acceso a detritus. Cocodrilos modernos pueden vivir como carroñeros y pueden sobrevivir durante meses sin comida, y sus jóvenes son pequeñas, crecen lentamente, y se alimentan en gran medida de los invertebrados y los organismos muertos o fragmentos de organismos para sus primeros años. Estas características se han relacionado con la supervivencia de los cocodrilos a finales del Cretácico.
Después del evento de extinción K-Pg, la biodiversidad requiere bastante tiempo para recuperarse, a pesar de la existencia de abundante vacante nichos ecológicos.
Microbiota
El K-T frontera representa uno de los volúmenes de negocios más dramáticos en el registro fósil para diversos calcáreo nanoplancton que formó las calcio depósitos que dieron el Cretácico su nombre. El volumen de negocios en este grupo está claramente marcada a nivel de especie. El análisis estadístico de pérdidas marinos en este momento sugiere que la disminución en la diversidad fue causada más por un fuerte aumento en las extinciones que por una disminución en especiación. El registro K-T límite de dinoflagelados no es tan bien entendidas, sobre todo porque sólo quistes microbianos proporcionar un registro fósil, y no todas las especies de dinoflagelados tienen etapas formadores de quistes, por lo tanto es probable que causa la diversidad para ser subestimada. Estudios recientes indican que no hubo grandes cambios en los dinoflagelados a través de la capa límite.
Radiolaria haber dejado un registro geológico desde al menos los Ordovícico veces, y sus esqueletos fósiles minerales puede ser rastreado a través del límite KT. No hay evidencia de extinción en masa de estos organismos, y hay un apoyo para alta productividad de estas especies en latitudes altas del sur como consecuencia de las temperaturas de enfriamiento a principios del Paleoceno. Aproximadamente el 46% de especies de diatomeas sobrevivieron a la transición desde el Cretácico superior al Paleoceno . Esto sugiere un volumen de negocios significativo en especies, pero no una extinción catastrófica de diatomeas, a través del límite KT.
La ocurrencia de planctónica foraminíferos a través del límite K-T ha sido estudiado desde los años 1930. Investigación estimulado por la posibilidad de un evento de impacto en el límite K-T resultó en numerosas publicaciones que detallan la extinción de foraminíferos planctónicos en el límite. Sin embargo, existe un debate en curso entre los grupos que creen que la evidencia indica extinción sustancial de estas especies en el límite KT, y los que creen que la evidencia apoya múltiples extinciones y expansiones a través de la frontera.
Numerosas especies de foraminíferos bentónicos se extinguió durante el evento de extinción K-Pg, presumiblemente porque dependen de los restos orgánicos de nutrientes, ya que el biomasa en el océano se cree que ha disminuido. Sin embargo, como la microbiota marina se recuperó, se cree que el aumento de la especiación de foraminíferos bentónicos como resultado del aumento de las fuentes de alimentos. El fitoplancton recuperación a principios del Paleoceno proporcionó la fuente de alimento para soportar grandes asociaciones de foraminíferos bentónicos, que son principalmente en periodo de detritus. La recuperación final de las poblaciones bentónicas ocurrió en varias etapas que duran varios cientos de miles de años en el Paleoceno.
Los invertebrados marinos

Hay una variabilidad en el registro fósil como a la tasa de extinción de invertebrados marinos en todo el límite KT. La tasa aparente está influenciada por la falta de registros fósiles en lugar de extinción real.
Ostrácodos, una clase de pequeños crustáceos que prevalecían en la parte superior del Maastrichtiano, dejó depósitos fósiles en una variedad de lugares. Una revisión de estos fósiles muestra que la diversidad ostrácodos fue menor en el Paleoceno que cualquier otro momento en la era terciaria. Sin embargo, la investigación actual no puede determinar si las extinciones ocurrieron antes o durante el propio intervalo límite.
Aproximadamente el 60% de finales de los Cretácico Scleractinia géneros de coral no pudo cruzar la frontera K-T en el Paleoceno. Un análisis más detallado de las extinciones de corales muestra que aproximadamente el 98% de las especies coloniales, los que habitan cálidas y poco profundas tropicales aguas, se extinguió. Los corales solitarios, que generalmente no forman arrecifes y habitan más frío y más profundo (por debajo de la zona) zonas fótica del océano se vieron menos afectados por el límite KT. Especies de corales coloniales dependen simbiosis con fotosintética de algas , que se derrumbó debido a los acontecimientos que rodearon el límite KT. Sin embargo, el uso de los datos a partir de fósiles de coral para apoyar la extinción y recuperación Paleoceno posterior K-Pg debe sopesarse frente a los cambios que se produjeron en los ecosistemas de coral a través del límite KT.
Los números de cefalópodo, equinodermos, y géneros bivalvo exhibió disminución significativa después de que el límite KT. La mayoría de las especies de braquiópodos, una pequeña phylum de invertebrados marinos, sobrevivió a la extinción K-Pg y diversificado durante el Paleoceno.

Excepto por nautiloides (representado por el orden moderno Nautilida) y Coleoideos (que ya tenían divergido en moderno octopodes, calamares, y sepia) todas las demás especies de la clase de moluscos Cephalopoda se extinguió en el límite KT. Éstos incluyeron la importancia ecológica belemnoids, así como los amonites , un grupo de muy diversos, numerosos y ampliamente distribuidos cefalópodos sin cáscara. Los investigadores han señalado que la estrategia reproductiva de los nautiloides supervivientes, que se basan en unos pocos y grandes huevos, desempeñó un papel en outsurviving sus homólogos sucesión con asociaciones a través del evento de extinción. Los amonoideos utilizan una estrategia planctónicas de reproducción (numerosos huevos y larvas planctónicas), que han sido devastadas por el evento de extinción K-Pg. Otras investigaciones han demostrado que después de esta eliminación de amonites de la biota mundial, nautiloides comenzó una radiación evolutiva en formas de concha y complejidades hasta entonces conocido sólo de amonites.
Aproximadamente el 35% de géneros equinodermo se extinguió en el límite KT, aunque taxones que prosperó en latitudes bajas, ambientes de aguas poco profundas durante el Cretácico tardío tuvo la tasa más alta de extinción. De latitud media, equinodermos de aguas profundas eran mucho menos afectados en el límite KT. El patrón de puntos de extinción a la pérdida de hábitat, específicamente el ahogamiento de plataformas carbonatadas, los arrecifes de aguas poco profundas en existencia en ese momento, por la extinción.
Otros grupos de invertebrados, incluyendo rudistas (almejas formadores de arrecifes) y inocerámidos (parientes gigantes de la moderna vieiras), también se extinguieron en el límite KT.
Pescado
Existen registros fósiles sustanciales de mandíbulas peces a través del límite K-T, que proporciona una buena evidencia de patrones de extinción de estas clases de vertebrados marinos. Dentro peces cartilaginosos, aproximadamente el 80% de los tiburones , rayos, y familias patines sobrevivieron al evento de extinción, y más de 90% de peces teleósteos (peces óseos) familias sobrevivieron. Hay pruebas de una matanza masiva de peces óseos en un yacimiento de fósiles inmediatamente por encima de la capa límite KT en Isla Seymour cerca de la Antártida, al parecer precipitada por el evento de extinción K-Pg. Sin embargo, los ambientes marinos y de agua dulce de peces mitigan los efectos ambientales de la extinción.
Invertebrados terrestres
Insectos daños a las hojas fosilizadas de plantas con flores de catorce sitios en América del Norte fueron utilizados como un indicador de la diversidad de insectos a través del límite K-T y se analizaron para determinar la tasa de extinción. Los investigadores encontraron que los sitios del Cretácico, antes del evento de extinción, tuvieron rica diversidad vegetal y los insectos que se alimentan. Sin embargo, a principios del Paleoceno, flora eran relativamente diversa con poca depredación de los insectos, incluso 1,7 millones años después del evento de extinción.
Las plantas terrestres
Hay pruebas abrumadoras de la interrupción mundial de las comunidades vegetales en el límite KT. Las extinciones se ven tanto en los estudios de polen fósil y hojas fósiles. En América del Norte, los datos sugieren una devastación masiva y la extinción masiva de plantas en las secciones límite KT, aunque hubo cambios sustanciales megafloral antes del límite. En América del Norte, aproximadamente el 57% de las especies vegetales se extinguieron. En latitudes altas del hemisferio sur, como Nueva Zelanda y la Antártida la masa mortandad de flora no causó ningún volumen de negocios significativo en especies, pero los cambios dramáticos y de corto plazo en la abundancia relativa de los grupos de plantas. En algunas regiones, la recuperación de las plantas Paleoceno comenzó con recolonizaciones por especies de helechos, representada como una pico de helecho en el registro geológico; Se observó el mismo patrón de recolonización helecho después del 1980 el Monte St. Helens erupción . Sin embargo, los patrones de recuperación eran bastante variable. Diferentes especies de helechos eran responsables de la espiga helecho en diferentes áreas, y en algunas regiones, sin pico helecho es evidente.
Debido a la destrucción total de las plantas en el límite KT hubo una proliferación de organismos saprotrophic tales como los hongos que no requieren de la fotosíntesis y utilizan los nutrientes de la vegetación en descomposición. El predominio de las especies de hongos duró sólo unos pocos años, mientras que el ambiente despejado y había un montón de materia orgánica para alimentarse. Una vez que la atmósfera se aclaró, los organismos fotosintéticos como los helechos y otras plantas devueltos. Polyploidy parece haber mejorado la capacidad de las plantas con flores para sobrevivir a la extinción, probablemente porque las copias adicionales del genoma de estas plantas poseían les permitió adaptarse más fácilmente a las condiciones ambientales cambian rápidamente que siguieron al impacto.
Anfibios
Hay pruebas limitadas para la extinción de los anfibios en el límite K-Pg. Un estudio de los vertebrados fósiles a través del límite K-Pg en Montana llegó a la conclusión de que ninguna de las especies de anfibios se extinguieron. Sin embargo, hay varias especies de anfibios Maastrichtiano, no incluidas como parte de este estudio, que son desconocido del Paleoceno. Estos incluyen la rana Theatonius lancensis y la albanerpetontid Albanerpeton Galaktion; Por lo tanto, algunos anfibios parecen haberse extinguido en el límite. Los niveles relativamente bajos de extinción visto entre los anfibios probablemente reflejan las bajas tasas de extinción observados en animales de agua dulce.
Reptiles no arcosaurios

Los dos no vive taxones de reptiles archosaurian, testudines (tortugas) y lepidosaurios ( serpientes, lagartos , y Amphisbaenia (lagartos gusano)), junto con choristoderes (semi-acuática archosauromorphs que se extinguió a principios del Mioceno ), sobrevivieron a través del límite KT. Más del 80% de las especies de tortugas del Cretácico pasaron por el límite KT. Además, las seis familias de tortugas en existencia al final del Cretácico sobrevivieron hasta el Terciario y están representados por las especies actuales. Lepidosaurios de vivienda incluye Rhynchocephalia ( tuataras) y Squamata. El Rhynchocephalia eran un grupo amplio y relativamente exitosa de lepidosaurios a principios del Mesozoico, pero comenzó a declinar por los mediados del Cretácico. Están representados hoy por un solo género ubicada exclusivamente en Nueva Zelanda.
El orden Squamata, que está representado hoy por lagartos, serpientes y Amphisbaenia, irradiaba en diversos nichos ecológicos durante el Jurásico y tuvieron éxito en todo el Cretácico. Sobrevivieron a través del límite KT y son actualmente el grupo más exitoso y diverso de reptiles que viven con más de 6.000 especies existentes. No conocida familia de los escamosos terrestres se extinguieron en el límite, y la evidencia fósil indica que no sufren ninguna disminución significativa en los números. Su pequeño tamaño, el metabolismo adaptable y capacidad de moverse a hábitats más favorables fueron factores clave en su supervivencia durante el Paleoceno tardío Cretáceo y principios. Reptiles acuáticos no archosaurian gigantes como mosasaurios y plesiosaurios, que eran los depredadores marinos superiores de su tiempo, se extinguieron a finales del Cretácico. Esto es más probable debido a su tamaño, lo que significaba que eran menos capaces de adaptarse durante las consecuencias de la extinción.
Arcosaurios
La clado arcosaurio incluye dos órdenes de vida, cocodrilos (de los cuales Alligatoridae, Crocodylidae y Gavialidae son las familias sólo sobreviven) y dinosaurios (de los cuales las aves son los únicos miembros supervivientes), junto con los no aviares extintos dinosaurios y pterosaurios .
Crocodyliformes
Diez familias de los cocodrilos o sus parientes cercanos son representados en los registros fósiles Maastrichtiano, de los cuales cinco murieron a cabo antes del límite KT. Cinco familias tienen dos representantes fósiles Maastrichtiano y Paleoceno. De agua dulce y terrestres entornos -excepto Todas las familias sobrevivientes de crocodiliformes habitadas por la Dyrosauridae, que vivió en agua dulce y localizaciones marinas. Aproximadamente el 50% de los representantes crocodyliforme sobrevivió a través del límite KT, la única tendencia aparente ser que no hay grandes cocodrilos sobrevivieron. Supervivencia crocodyliforme través de la frontera puede haber resultado de su nicho acuático y la capacidad de excavar, lo que reduce la susceptibilidad a los efectos ambientales negativos en el límite. Jouve y sus colegas sugirieron en 2008 que crocodiliformes juvenil marinos vivían en ambientes de agua dulce como modernos juveniles cocodrilo marino, lo que habría ayudado a sobrevivir donde otros reptiles marinos se extinguieron; ambientes de agua dulce no estaban tan fuertemente afectados por el evento de extinción K-Pg como ambientes marinos.
Los pterosaurios
Una familia de pterosaurios , Azhdarchidae, fue sin duda presente en el Maastrichtiano, y se extinguió en el límite KT. Estas grandes pterosaurios fueron los últimos representantes de un grupo decreciente que contenía 10 familias durante los mediados del Cretácico. Pterosaurios más pequeños se extinguieron antes de la Maastrichtiano durante un período que vio una disminución de las especies animales más pequeños, mientras que las especies más grandes se hicieron más frecuentes. Recientemente, varios taxones de pterosaurios se han descubierto data del Campaniano / Maastrichtiano, como el ornithocheirids Piksi y " Ornithocheirus ", posible pteranodontids y nyctosaurids, y una tapejarid. Mientras esto ocurre, aves modernas fueron sometidos a la diversificación y la sustitución de las aves arcaicas y grupos de pterosaurios, posiblemente debido a la competencia directa, o simplemente llenan nichos vacíos.
Aves
La mayoría de los paleontólogos miran pájaros como los dinosaurios sólo sobrevivieron (véase Origen de las aves). Sin embargo, no todos aves neornithean se extinguieron, incluyendo grupos florecientes como enantiornithines y hesperornithiforms. Varios análisis de fósiles de aves muestran divergencia de especies antes del límite KT, y que el pato, pollo y parientes de aves de ratites coexistieron con los dinosaurios no aviares. Las grandes colecciones de fósiles de aves que representan una gama de diferentes especies proporciona evidencia definitiva de la persistencia de las aves arcaicas dentro de 300.000 años del límite K-Pg. Ninguno de ellos son conocidos por sobrevivir en el Paleógeno, y su persistencia en la última Maastrichtiano por lo tanto proporciona una fuerte evidencia de una extinción masiva de aves arcaicas coincidiendo con el impacto de un asteroide de Chicxulub. Una pequeña fracción de las especies de aves cretácicos sobrevivió el impacto, dando lugar a las aves actuales. Hasta ahora, sólo una única especie de aves, que no ha sido nombrado, se ha identificado con confianza tanto por encima como por debajo del límite K-Pg (que está presente en el Maastrichtiano Formación Hell Creek y Danian Fort Union Formación). El único grupo de aves conocido por seguro que han sobrevivido a la K-T es el límite Neornithines (aunque una especie Paleógeno, Qinornis paleocenica, puede representar un no neornithine pájaro sobreviviente). Neornithines pueden haber sido capaz de sobrevivir a la extinción como resultado de sus habilidades para bucear, nadar, o buscar refugio en el agua y pantanos. Muchas especies de neornithines pueden construir madrigueras o nidos en huecos de los árboles o nidos de termitas, todos los cuales proporcionan refugio de los efectos ambientales en el límite KT. La supervivencia a largo plazo más allá de la frontera fue asegurado como consecuencia de llenar nichos ecológicos vacíos dejados por la extinción de los dinosaurios no aviares.
Dinosaurios no aviares

Excluyendo algunas reclamaciones controversiales, los científicos están de acuerdo en que todos los dinosaurios no aviares se extinguieron en el límite KT. El registro fósil de dinosaurio ha sido interpretado para mostrar tanto una disminución de la diversidad y la no disminución de la diversidad en los últimos millones de años del Cretácico, y puede ser que la calidad del registro fósil de dinosaurio no es simplemente lo suficientemente bueno como para permitir a los investigadores distinguir entre las opciones. Dado que no hay evidencia de que los dinosaurios no aviares a finales del Maastrichtiano podrían excavar, nadar o bucear, no fueron capaces de protegerse de las peores partes de ningún tipo de estrés ambiental que se produjo en el límite KT. Es posible que los pequeños dinosaurios (que no sean aves) sobrevivieron, pero habría sido privado de alimentos ya que ambos dinosaurios herbívoros habrían encontrado material vegetal escasa y carnívoros habría encontrado rápidamente presas escasean.
El creciente consenso acerca de la endotermia de los dinosaurios (ver fisiología de los dinosaurios) ayuda a comprender su plena extinción en contraste con sus parientes cercanos, el cocodrilos. Ectotérmicos («a sangre fría") cocodrilos tienen necesidades muy limitadas de alimentos (que pueden sobrevivir varios meses sin comer) mientras ("sangre-caliente") animales endotérmicos de tamaño similar necesitan mucha más comida para sostener su metabolismo más rápido. Por lo tanto, en las circunstancias de interrupción de la cadena alimentaria se ha mencionado anteriormente, los dinosaurios no aviares murieron, mientras que algunos cocodrilos sobrevivieron. En este contexto, la supervivencia de otros animales endotérmicos, como algunas aves y mamíferos, podría deberse, entre otras razones, a sus necesidades más pequeñas de alimentos, en relación con su tamaño pequeño en la época extinción.
Si la extinción se produjo gradualmente o de repente se ha debatido, ya que ambos puntos de vista tienen apoyo en el registro fósil. Un estudio de 29 yacimientos de fósiles en catalán Pirineo de Europa en 2010 apoya que los dinosaurios allí tenían gran diversidad hasta que el impacto de un asteroide. Otros han interpretado las rocas fosilíferas largo Río Red Deer en Alberta, Canadá, el apoyo a una paulatina extinción de los dinosaurios no aviares como; durante los últimos 10 millones de años de las capas del Cretácico allí, el número de especies de dinosaurios parece haber disminuido de 45 a 12. Otros científicos han señalado la misma.
Varios investigadores apoyan la existencia de Dinosaurios del Paleoceno. Evidencia de esta existencia se basa en el descubrimiento de restos de dinosaurio en el Formación Hell Creek de hasta 1,3 m (4,3 pies) por encima y 40.000 años más tarde que el límite KT. Muestras de polen se recuperaron cerca de un fósil hadrosaurio fémur recupera en la Arenisca de Ojo Álamo en el Río San Juan indican que el animal vivió durante el Terciario, aproximadamente 64.5 Ma (aproximadamente 1 millón de años después de la extinción K-Pg). Si su existencia más allá de la K-T límite puede confirmarse, estos hadrosáuridos sería considerado un Walking Dead Clade. El consenso científico es que estos fósiles fueron erosionadas de sus ubicaciones originales y luego re-enterrados en sedimentos mucho más tarde (también conocidos como fósiles reelaborados).

Mamíferos
Todos los principales Cretácico mamíferos linajes, incluyendo monotremas (mamíferos que ponen huevos), multituberculados, marsupiales y placentarios, dryolestoideans, y gondwanatheres sobrevivió a la extinción K-Pg, a pesar de que sufrieron pérdidas. En particular, los marsupiales desaparecieron en gran parte de América del Norte y Asia deltatheroidans, parientes primitivos de marsupiales existentes, se extinguieron. En las camas de Hell Creek de América del Norte, al menos la mitad de las diez especies de multituberculados conocidos y las once especies de marsupiales no se encuentran por encima del límite.
Especies de mamíferos comenzaron a diversificar aproximadamente 30 millones de años anteriores al límite KT. La diversificación de los mamíferos se estancó a través del límite. Las investigaciones actuales indican que los mamíferos no se diversificaron explosivamente a través del K-T frontera, a pesar de los nichos ambientales proporcionados por la extinción de los dinosaurios. Varias órdenes de mamíferos han sido interpretadas como diversificar inmediatamente después de la K-T frontera, incluyendo quirópteros (murciélagos) y Cetartiodactyla (un grupo diverso que incluye hoy ballenas y delfines y ungulados incluso con los dedos), aunque la investigación reciente concluye que sólo órdenes de marsupiales diversificaron después del límite KT.
K-T especies de mamíferos de contorno eran generalmente pequeña, comparable en tamaño a las ratas ; este pequeño tamaño les habría ayudado a encontrar refugio en ambientes protegidos. Además, se postula que algunos de los primeros monotremas, marsupiales y placentarios, eran semiacuática o madriguera, ya que hay múltiples linajes de mamíferos con tales hábitos hoy en día. Cualquier madriguera o mamífero semiacuático habrían tenido una protección adicional contra K-T tensiones ambientales transfronterizos.
Evidencia
Fósiles de América del Norte
En secuencias terrestres de América del Norte, el evento de extinción está mejor representado por la marcada discrepancia entre los ricos y relativamente abundantes de finales del Maastrichtiano registro de palinomorfos y la espiga helecho post-límite.
En la actualidad, la secuencia más informativo de las rocas de dinosaurios que devengan en el mundo desde el límite KT se encuentra en el oeste de América del Norte, sobre todo a finales del Edad Maastrichtiano Formación Hell Creek de Montana, Estados Unidos. Esta formación, en comparación con el antiguo (aproximadamente 75 Ma) Río Judith / Parque de los Dinosaurios Formaciones (de Montana, EE.UU., y Alberta, Canadá, respectivamente) proporciona información sobre los cambios en las poblaciones de dinosaurios en los últimos 10 millones de años de los Cretácico. Estas camas fósiles son geográficamente limitada, que cubre sólo una parte de un continente.
Las formaciones de la Campania de mediana finales muestran una mayor diversidad de dinosaurios que cualquier otro grupo de rocas.Los fines de rocas Maastrichtiano contienen los miembros más grandes de varios clados principales:Tyrannosaurus,Ankylosaurus,Pachycephalosaurus,Triceratopsy Torosaurus, lo que sugiere la comida era abundante inmediatamente antes de la extinción.
Además de ricas fósiles de dinosaurios, también hay fósiles de plantas que ilustran la reducción de las especies de plantas a través del límite K-T. En los sedimentos debajo del límite KT los restos vegetales dominantes son los granos de polen de angiospermas, pero la capa límite real contiene poco polen y está dominado por las esporas de helechos. Niveles de polen normales reanudar gradualmente por encima de la capa límite. Esto es una reminiscencia de las zonas asoladas por las erupciones volcánicas modernas, donde la recuperación está dirigido por helechos, que luego son reemplazados por las plantas angiospermas más grandes.
Fósiles marinos
La extinción masiva de plancton marino parece haber sido brusco y justo en el límite KT. amonita géneros se extinguió en o cerca del límite KT; sin embargo, hubo una extinción más pequeño y más lento de los géneros ammonite antes de la frontera que se asoció con una regresión marina cretáceo tarde. La extinción gradual de la mayoría de los bivalvos inocerámidos comenzó mucho antes del límite KT, y una pequeña reducción, gradual en la diversidad de la amonita se produjo a través de las muy tardías Cretácico. Un análisis más detallado muestra que varios procesos se encontraban en curso a finales de los mares del Cretácico y parcialmente se superponen en el tiempo, y luego terminó con la extinción masiva abrupta.
Megatsunamis
El consenso científico es que el impacto bólido en los K-T frontera dejado depósitos de tsunami y sedimentos alrededor de la zona del Mar Caribe y el Golfo de México. Estos depósitos se han identificado en la cuenca de La Popa en el noreste de México , carbonatos de plataforma en el noreste de Brasil, y los sedimentos de aguas profundas del Atlántico.
Duración
La cantidad de tiempo necesario para que la extinción se produzca es un tema controvertido, ya que algunas teorías sobre las causas de la extinción requieren una rápida extinción en un período relativamente corto (de unos años a unos pocos miles de años), mientras que otros requieren períodos más largos. El problema es difícil de resolver debido al efecto Signor-Lipps; es decir, el registro fósil es tan incompleta que especies más extintos probablemente se extinguieron poco después de que el fósil más reciente que se ha encontrado. Los científicos también han encontrado muy pocas camas continuas de roca que contiene fósiles que cubren un rango de tiempo de varios millones de años antes de la extinción K-Pg de unos pocos millones de años después de la misma.
Impacto de un asteroide de Chicxulub


La evidencia de impacto
En 1980, un equipo de investigadores que consisten en premio Nobel el físico Luis Álvarez, su hijo geólogo Walter Álvarez, y los químicos Frank Asaro y Helen Michel descubrió que sedimentarias capas encontrar en todo el mundo en el límite Cretácico-Paleógeno contienen una concentración de iridio muchas veces mayor de lo normal (30, 160 y 20 veces en tres secciones estudiadas originalmente). Iridium es extremadamente raro en la corteza de la Tierra , porque es un elemento siderophile, y por lo tanto la mayor parte de ella viajó con el hierro , ya que se hundió en el núcleo de la Tierra durante diferenciación planetaria. Como iridio sigue siendo abundante en la mayoría de los asteroides y cometas , el equipo de Álvarez sugirió que un asteroide golpeó la Tierra en el momento del límite K-T. Hubo especulaciones anteriores sobre la posibilidad de un evento de impacto, pero esta fue la primera evidencia sólida de un impacto.
Esta hipótesis fue visto como radical cuando se propuso por primera vez, pero pronto surgió evidencia adicional. La arcilla límite se encontró a estar lleno de esférulas minutos de roca, cristalizados de gotitas de roca fundida que se forma durante el impacto. El cuarzo impactado y otros minerales también se identificaron en el límite K-Pg. Impresionado minerales tienen su estructura interna deforma y son creados por intensas presiones tales como los asociados con explosiones nucleares o impactos de meteoritos. La identificación de las camas gigantes del tsunami a lo largo de la costa del Golfo y el Caribe también proporcionó evidencia de impacto, y sugirió que el impacto pudo haber ocurrido cerca- como lo hizo el hecho de que el límite K-Pg convirtió más gruesa en el sur de Estados Unidos, con metros- camas gruesas de los desechos que se producen en el norte de Nuevo México.
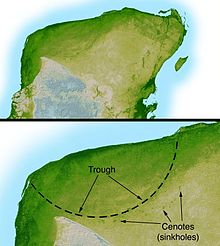
Más investigación identificó el cráter Chicxulub gigante, enterrada bajo Chicxulub en la costa de Yucatán, México como la fuente de la arcilla límite K-Pg. Identificado en 1990 con base en el trabajo de Glen Penfield 1978, el cráter es ovalada, con un diámetro medio de alrededor de 180 kilómetros (110 millas), aproximadamente del tamaño calculado por el equipo de Álvarez. El descubrimiento del Cráter una predicción necesaria del impacto de hipótesis aportan pruebas concluyentes de un impacto K-Pg, y reforzó la hipótesis de que la extinción fue causada por un impacto.
En 2007, una hipótesis fue planteada que argumentó el impactador que mataron a los dinosaurios pertenecían a la familia de asteroides Baptistina. Se han planteado preocupaciones con respecto a la relación de renombre, en parte porque existen muy pocas restricciones observacionales sólidos del asteroide o familiar. De hecho, se descubrió recientemente que 298 Baptistina no comparte la misma firma química como la fuente del impacto K-Pg. Aunque este hallazgo puede hacer que el vínculo entre la familia Baptistina y K-Pg impactador más difícil de justificar, no se opone a la posibilidad. Un 2011 estudio WISE de la luz reflejada por los asteroides de la familia estima la ruptura a 80 Ma, dándole tiempo suficiente para cambiar de órbitas e impactar la Tierra en un 66 Ma.
Paul Renne del Centro de Geocronología de Berkeley ha informado de que la fecha del evento asteroide es de hace 66.038 millones años, más o menos 11.000 años, basado en la desintegración radiactiva de argón . Se postula además que la extinción masiva ocurrió dentro de 33.000 años de esta fecha.
Efectos de impacto
Tal impacto habría inhibido la fotosíntesis mediante la generación de una nube de polvo que bloqueó la luz del sol durante un año o menos, y mediante la inyección de ácido sulfúrico aerosoles en la estratosfera, que habría reducido la luz solar llega a la superficie de la Tierra en un 10-20%. Se necesitarían al menos diez años para los aerosoles a disiparse, lo que explicaría la extinción de plantas y fitoplancton, y de los organismos que dependen de ellos (incluidos los animales depredadores, así como los herbívoros ). Las pequeñas criaturas cuyas cadenas de alimentos se basaban en detritus tendrían una posibilidad razonable de supervivencia. Las consecuencias de la reentrada de material expulsado a la atmósfera de la Tierra incluirían unas breves (horas largas) pero intenso pulso de radiación infrarroja, matando a los organismos expuestos. Global tormentas de fuego pueden ser consecuencia del pulso de calor y la caída a la Tierra de fragmentos incendiarios de la explosión. Los altos O 2 niveles durante el Cretácico tardío habrían apoyado la combustión intensa. El nivel de la atmósfera O 2 se desplomó a principios del período terciario. Si se han producido incendios generalizados, habrían aumentado el CO 2 contenido de la atmósfera y provocó un temporal efecto invernadero una vez que la nube de polvo se asentó, y esto habría exterminado a los organismos más vulnerables que sobrevivieron al período inmediatamente después del impacto.
El impacto también puede haber producido la lluvia ácida , dependiendo de qué tipo de roca golpeó el asteroide. Sin embargo, la investigación reciente sugiere que este efecto fue relativamente menor, con una duración de aproximadamente 12 años. La acidez fue neutralizado por el medio ambiente y la supervivencia de los animales vulnerables a los efectos de la lluvia ácida (como ranas ) indican que esto no era un importante contribuyente a la extinción. Teorías de impacto sólo pueden explicar las extinciones muy rápidos, ya que las nubes de polvo y posibles aerosoles sulfúricos se lave fuera de la atmósfera en un tiempo bastante corto, posiblemente menos de diez años.
La forma y la ubicación del cráter indican otras causas de la devastación además de la nube de polvo. El asteroide caído en el océano y tendría causados megatsunamis, para el que se ha encontrado evidencia en varios lugares en la arena Estados Unidos-marina del Caribe y del este en lugares que estaban entonces en el interior, y restos de vegetación y rocas terrestres en sedimentos marinos de fecha a la momento del impacto. El asteroide cayó en un lecho de yeso ( sulfato de calcio), lo que habría producido una vasta dióxido de azufre en aerosol. Esto tendría reduce aún más la luz solar llega a la superficie de la Tierra y luego se precipita en forma de lluvia ácida, la vegetación matar, plancton y organismos que construyen conchas de carbonato de calcio ( cocolitóforos y moluscos). En febrero de 2008, un equipo de investigadores utiliza imágenes sísmicas del cráter para determinar que el impactador aterrizó en aguas más profundas de lo que se suponía anteriormente. Se argumenta que esta habría dado lugar a un aumento de los aerosoles de sulfato en la atmósfera, lo que podría haber hecho el impacto más letal mediante la alteración del clima y mediante la generación de lluvia ácida.
La mayoría de los paleontólogos están de acuerdo en que un asteroide golpeó la Tierra aproximadamente al final del Cretácico, pero hay una disputa si el impacto fue la única causa de las extinciones. Hay pruebas de que hubo un intervalo de alrededor de 300 ka del impacto de la extinción masiva. En 1997, el paleontólogo Sankar Chatterjee señaló la propuso y mucho más grande 600 kilometros (370 millas) Shiva cráter y la posibilidad de un escenario de múltiples impactos.
En marzo de 2010 un grupo internacional de científicos hizo suya la hipótesis de asteroides, específicamente el impacto de Chicxulub, como la causa de la extinción. Un equipo de 41 científicos revisó 20 años de literatura científica y, al hacerlo, también descartó otras teorías, como el vulcanismo masivo. Se habían determinado que un 10 a 15 km (09.06 mi) roca espacial se precipitó hacia la Tierra en Chicxulub en México de Península de Yucatán. La colisión habría lanzado la misma energía que 100 teratonnes de TNT (420 ZJ), más de mil millones de veces la energía de la bombas lanzadas sobre Hiroshima y Nagasaki.
Las hipótesis alternativas
El hecho de que las extinciones se producen al mismo tiempo que el impacto de un asteroide Chicxulub apoya firmemente la hipótesis de impacto de la extinción. Sin embargo, algunos científicos siguen disputar el papel del impacto de Chicxulub en la conducción de la extinción, y sugerir que otros eventos pueden haber contribuido a la extinción masiva del Cretácico-end. En particular, las erupciones volcánicas, el cambio climático, el cambio del nivel del mar, y otros eventos de impacto se han propuesto para desempeñar un papel en el impulso de la extinción K-Pg.
Trampas del Decán
Antes de 2000, los argumentos que las Deccan Traps basaltos de inundación causaron la extinción fueron por lo general vinculados a la opinión de que la extinción fue gradual, ya que se pensaba que los acontecimientos de la inundación de basalto haber comenzado alrededor de 68 Ma y duraron por más de 2 millones de años. La evidencia más reciente muestra que las trampas estallaron más de 800.000 años que abarca el límite K-T, y por lo tanto puede ser responsable de la extinción y la recuperación biótica retardada a partir de entonces.
Las Trampas del Decán podría haber causado la extinción a través de varios mecanismos, incluyendo la liberación de polvo y aerosoles sulfúricos en el aire, lo que podría haber bloqueado la luz solar y por lo tanto reduce la fotosíntesis en las plantas. Además, Deccan Trampa vulcanismo podría haber dado lugar a las emisiones de dióxido de carbono que incrementaron el efecto invernadero cuando el polvo y aerosoles eliminan de la atmósfera.
En los años en que la hipótesis de Deccan Traps fue vinculado a una extinción más lenta, Luis Alvarez (que murió en 1988) respondió que los paleontólogos estaban siendo engañados por la escasez de datos. Aunque no fue inicialmente bien recibida su afirmación, los estudios de campo más tarde intensivas de yacimientos de fósiles prestaron peso a su reclamación. Con el tiempo, la mayoría de los paleontólogos comenzaron a aceptar la idea de que las extinciones masivas al final del Cretácico eran en gran parte o por lo menos en parte debido a un impacto Tierra masiva. Sin embargo, incluso Walter Álvarez ha reconocido que hubo otros cambios importantes en la Tierra, incluso antes del impacto, como una caída en el nivel del mar y las erupciones volcánicas masivas que produjeron las Trampas de Deccan de la India, y estos pueden haber contribuido a la extinción.
Evento de impacto múltiple
Varios otros cráteres también parecen haberse formado sobre el tiempo del límite K-T. Esto sugiere la posibilidad de múltiples impactos simultáneos cerca, tal vez de un objeto asteroidal fragmentada, similar al impacto Shoemaker-Levy 9 con Júpiter . Además de los 180 km (110 millas) Cráter de Chicxulub, hay 24 km (15 millas) del cráter Boltysh en Ucrania (65.17 ± 0.64 Ma) , los 20 km (12 millas) del cráter Silverpit, un cráter de impacto de sospecha en el Norte Mar (60-65 Ma) , y el controvertido y mucho más grande 600 kilometros (370 millas) Shiva cráter. Cualesquiera otros cráteres que podrían haberse formado en el océano Tetis habrían sido oscurecida por eventos tectónicos como la deriva hacia el norte implacable de África y la India.
Regresión Maastrichtiano del nivel del mar
Existe una clara evidencia de que los niveles del mar cayeron en la etapa final del Cretácico en más que en cualquier otro momento de los Era Mesozoica. En algunos Maastrichtiano capas etapa roca de varias partes del mundo, las capas posteriores son terrestres; capas anteriores representan los litorales y las primeras capas representan los fondos marinos. Estas capas no muestran la inclinación y la distorsión asociada con la formación de montañas, por lo tanto, la explicación más probable es una "regresión", es decir, una caída en el nivel del mar. No hay evidencia directa de la causa de la regresión, pero la explicación aceptada actualmente como más probable es que las dorsales oceánicas se volvieron menos activos y por lo tanto se hundieron bajo su propio peso.
Una regresión severa tendría reduce en gran medida el área de la plataforma continental, que es el más rico en especies parte del mar, por lo que podría haber sido suficiente para causar una marina extinción masiva. Sin embargo la investigación concluye que este cambio habría sido insuficiente para hacer que el nivel observado de la extinción de la amonita. La regresión también habría causado cambios climáticos, en parte mediante la interrupción de los vientos y las corrientes oceánicas y en parte por la reducción de la Tierra albedo y por lo tanto el aumento de las temperaturas globales.
Regresión marina también dio lugar a la pérdida de los mares epeiric, como el Mar interior occidental de América del Norte. La pérdida de estos mares alterado en gran medida los hábitats, la eliminación de las llanuras costeras que diez millones de años antes habían sido sede de diversas comunidades, como se encuentran en rocas de la Formación Dinosaur Park. Otra consecuencia fue una expansión de ambientes de agua dulce, ya que la escorrentía continental ahora tenía largas distancias para viajar antes de llegar a los océanos. Si bien este cambio fue favorable a agua dulce vertebrados , los que prefieren marinos entornos, como los tiburones , sufrieron.
Múltiples causas
En un artículo de revisión, J. David Archibald y David E. Fastovsky discutieron un escenario que combina tres causas principales postulados: vulcanismo, regresión marina y de impacto extraterrestre. En este escenario, las comunidades terrestres y marinos se destacaron por los cambios y la pérdida de hábitats. Los dinosaurios, como los vertebrados más grandes, fueron los primeros afectados por los cambios ambientales, así como su diversidad declinaron. Al mismo tiempo, materiales particulados de vulcanismo áreas del planeta se enfrió y se secan. Entonces, ocurrió un evento de impacto, provocando colapsos en las cadenas de comida basada en la fotosíntesis, tanto en las cadenas alimenticias terrestres ya estresados y en las cadenas alimentarias marinas. La principal diferencia entre esta hipótesis y las hipótesis de una sola causa es que sus defensores consideran las causas individuales propuestos, ya sea como no suficiente fuerza para causar la extinción o no susceptibles de producir el modelo taxonómico de la extinción.
Recuperación y radiación
La extinción K-Pg tuvo un profundo efecto sobre la evolución de la vida en la tierra. La eliminación de los grupos dominantes del Cretácico permitió a otros organismos para tomar su lugar, estimulando una notable serie de radiaciones adaptativas en el Paleógeno. El ejemplo más llamativo es la sustitución de los dinosaurios por los mamíferos. Después de la extinción K-Pg, los mamíferos evolucionaron rápidamente para llenar los nichos dejados vacantes por los dinosaurios. Dentro de los géneros de mamíferos, nuevas especies fueron aproximadamente 9,1% más grande después del límite KT.
Otros grupos también fueron sometidos a importantes radiaciones. En base a la secuenciación molecular y citas fósiles, Neoaves parecía irradiar después de que el límite KT. Incluso producen gigante, formas voladoras, como los herbívoros Gastornis y dromornithidae, y el depredador Phorusrhacidae. La extinción del Cretácico lagartos y serpientes puede haber dado lugar a la radiación de los grupos modernos tales como iguanas, lagartos y boas. En tierra, boid gigante y enormes serpientes madtsoiid aparecieron, y en los mares, gigantescas serpientes marinas radiadas. Peces teleósteos diversificó de forma explosiva, llenando los nichos dejados vacantes por la extinción. Grupos que aparecen en el Paleoceno y Eoceno incluyen marlines, atunes, anguilas y peces planos. Los principales cambios se observan también en las comunidades de insectos Paleógeno. Muchos grupos de hormigas estaban presentes en el Cretácico, pero en las hormigas del Eoceno se volvieron dominantes y diversa, con las colonias más grandes. Mariposas diversificados, así, tal vez para tomar el lugar de los insectos comedores de hojas exterminados por la extinción. Las avanzadas termitero, Termitidae, también subieron a la prominencia.