
{kind=link}
Vision humaine
La vision humaine est la perception humaine des rayonnements lumineux, c'est-à-dire, l'interprétation cognitive du sens de la vue, partagé par de nombreuses espèces animales.
La vision recouvre l'ensemble des processus physiologiques et psychologiques par lesquels la lumière émise ou réfléchie par l'environnement détermine les détails des représentations sensorielles, comme les formes, les couleurs, les textures, le mouvement, la distance et le relief. Ces processus processus cognitifs complexes font intervenir l'œil, organe récepteur de la vue, mais aussi des zones spécialisées du cerveau (voir cortex visuel).
Le système visuel
Le flux d'information en provenance de l'extérieur détecté par la rétine de l'œil n'est pas le seul facteur rentrant en compte dans la vision. Les illusions d'optique en sont la preuve la plus élémentaire : elles montrent que la façon dont le système visuel, et en particulier le cortex visuel du cerveau, traite cette information est aussi importante dans la construction de l'image perçue, consciemment ou non.
La vision n'est ni instantanée ni fluide, mais elle se fait de manière ponctuelle et rapide (de l'ordre du 1/40 de seconde). Le train d'informations visuelles passe depuis la rétine par les nerfs optiques pour être acheminé vers les aires corticales de la vision à l'arrière du cerveau. La façon dont le cerveau traite ces informations fait l'objet de nombreuses études en neurosciences cognitives, notamment depuis les travaux des Prix Nobel Hubel et Wiesel.
Au sein du système visuel, il a été décrit de nombreuses voies qui forment une architecture complexe chargée de traiter les informations de forme, le mouvement, l'identification des objets, la perception des visages, etc. Ainsi, par exemple, la sensation de relief n'est perçue qu'au travers de la vision combinée des deux yeux, traitée pour cela par le cerveau qui reconstitue le relief à partir de deux images légèrement décalées. Ce phénomène est exploité par la technique de la stéréoscopie.
Pour l'homme, ainsi que pour de nombreux animaux, la vision diffère selon l'intensité lumineuse. Il est d'usage de distinguer la vision diurne, ou « vision photopique », la vision nocturne, ou « vision scotopique », et le domaine intermédiaire ou « mésopique ». L'acuité visuelle est la finesse de la vision diurne des détails.
-

Courbes d'absorbance des cônes et des bâtonnets.
-

Comparaison des courbes de sensibilités relatives en vision photopique et scotopique
-
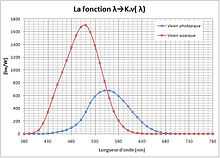
Sensibilité visuelle
L'espèce humaine perçoit un spectre lumineux de rayonnements électromagnétiques de longueur d'onde dans le vide [n 1]comprise environ entre 380 et 780 nanomètres (nm).
La sensibilité visuelle est différente dans la région centrale de l'œil dans des conditions d'éclairement suffisantes, avec perception des formes, des détails et des couleurs, et dans la région périphérique, ou dans des conditions lumineuses insuffisantes, avec une détection du mouvement principalement et sans vision des couleurs.
Vision photopique
Quand l'éclairement est suffisant, la répartition spectrale de ces rayonnements donne lieu à une sensation de couleur.

Chez l'homme, la vision photopique ou maculaire (macula) s'effectue, dans la partie centrale de l'œil exclusivement, grâce aux 5 à 7 millions de cônes de la rétine de chaque œil, répartis en trois types sensibles à des énergies photoniques différentes. La sensibilité des cônes est déterminée par des pigments dont un rayonnement d'énergie appropriée provoque la destruction, avec émission d'un influx nerveux ; l'organisme reconstitue continument les pigments avec un apport d'énergie.
Les sensibilités des types L et M, en léger décalage, concernent les énergies les plus faibles, correspondant aux courtes longueurs d'ondes. La concordance de leur signaux donne lieu à la perception de la luminosité ; la différence permet de distinguer le rouge du vert. La sensibilité du type S est nettement moindre, et concerne les photos à plus grande énergie. La différence entre les signaux des cônes S et celle des cônes L et M permet de distinguer la proportion de lumières bleues dans ce qu'on voit[n 2].
Les pigments des cônes L et M sont des protéines codées génétiquement sur le chromosome X, celle des cônes S sur le chromosome 7. Cette vision trichromique est rare ; parmi les mammifères, elle n'existe que chez certains primates[1].
La sensibilité photopique varie avec la fréquence des rayonnements. La sensibilité maximale correspond à un rayonnement de 555 nm. Une lumière où un tel rayonnement domine se perçoit d'une couleur vert-jaune.
Les différences d'influx entre les cônes sont transformés, dans des cellules nerveuses spécialisées de l'œil, en informations visuelles.
L'information transmise par les différents cônes est intégrée à différents niveaux des voies visuelles depuis les systèmes rétiniens d'opposition de couleur jusqu'au cortex visuel, en particulier l'aire V4. Le fonctionnement du système visuel est complexe.
Anomalies de la vision des couleurs
Une anomalie génétique conduit à une vision des couleurs réduite chez les personnes atteintes par le daltonisme. Lorsqu'aucun cône ne fonctionne, l'individu ne voit aucune couleur, et on dit alors qu'il est atteint d'achromatopsie.
Une mutation rare conduisant à une vision tétrachromique a été mise en évidence chez quelques femmes[2]. Aux trois types de cônes communs s'ajoute un quatrième, dont la sensibilité est décalée de 4 à 7 nm par rapport aux pigments M et L. Il semble que ces sujets ne perçoivent pas les couleurs différemment des humains trichromates[3]. Cette particularité ne change pas en effet la perception des couleurs, mais pourrait amener à des différences de métamérisme, les sujets voyant différentes des couleurs jugées identiques par les autres.
Visions scotopique et périphérique
La vision scotopique et la vision périphérique, s'effectuent principalement grâce aux bâtonnets de la rétine, beaucoup plus sensibles que les cônes ; ils ne permettent pas, à eux seuls, de distinguer les couleurs, et leur sensibilité maximale correspond à un rayonnement d'environ 510 nm (vert). C'est une vision très adaptée à la pénombre, contrairement à la vision maculaire nécessitant une intensité lumineuse élevée. Il y a peu de bâtonnets au centre de la rétine (il n'y en a même quasiment pas au niveau de la fovéa), ce qui fait que si l'on regarde directement un objet peu lumineux de nuit, il se peut qu'on ne le voie pas alors qu'on peut le voir si l'on regarde un peu à côté. Le pigment photosensible (rhodopsine) des cellules en bâtonnet met beaucoup plus de temps à se reconstituer après un éblouissement que les pigments des cellules en cône (jusqu'à plusieurs dizaines de minutes pour un rétablissement parfait).
On a montré[4] (chez le macaque dans un premier temps, et via la mesure de l'activité électrique neuronale) que quand un individu tourne le regard sur le côté, son cerveau adapte immédiatement le traitement des informations transmises par l'œil, en mobilisant les neurones de la vision périphérique, qui sont alors au maximum de leur activité. Ainsi le cerveau de quelqu'un qui marche en regardant de côté reste alerté quant aux risques de collision avec un objet situé devant lui.
Ce fait offre de nouvelles pistes pour la rééducation de patients souffrant de dégénérescence maculaire incurable, mais dont la vision périphérique est conservée.
Perception du mouvement
Le système visuel n'est sensible qu'au mouvement ; du point de vue de l'utilité, c'est la seule chose qui compte pour les êtres vivants, aussi la sélection naturelle a privilégié les êtres capables de le détecter. Pour regarder un objet fixe, nous bougeons sans cesse les yeux, et les partie cérébrales du système visuel, reliant les mouvements de l'image rétinienne à ceux des yeux, construisent les formes. La vision périphérique est sensible exclusivement au mouvement[5].
On a longtemps cru que le cinéma se servait simplement de la persistance rétinienne pour donner l'illusion du mouvement[réf. souhaitée]. En réalité le mouvement observé sur un écran semble essentiellement être une création du cerveau. On distingue quatre phénomènes dans cette illusion :
- Effet de continuité créé par la succession rapide des images (12 images par seconde pour les films d'animation et 18 images par seconde pour les films muets - passés à 24 images par seconde avec le cinéma sonore uniquement pour permettre une intelligibilité suffisante de la bande son). Mais le mouvement n'a l'air tout à fait fluide que vers 50 images par seconde. Par exemple, quand au cinéma il y a un panoramique assez rapide, on peut percevoir que le mouvement est saccadé, ce qui reflète la succession des images. Cela est aussi dû au fait que l'obturateur s'ouvre et se ferme 48 fois par seconde, ce qui signifie que chaque image est présentée deux fois, cela pour éviter le papillotement ou scintillement[réf. souhaitée].
- Les premiers films de synthèse, dont chaque image était nette, créaient une impression peu naturelle. On s'aperçut vers 1980 que l'introduction d'un flou artificiel proportionnel au mouvement, comme sur une "vraie" pellicule, donnait paradoxalement un effet plus réaliste[réf. souhaitée].
- Disparition du scintillement. On obtient cet effet en vision centrale vers 50 images par seconde. C'est le cas, par exemple, de la télévision à tube cathodique et à affichage entrelacé (2 x 25 ou 2 x 30 images par seconde). Mais si l'on regarde en vision périphérique (il suffit de regarder à côté de l'écran tout en portant son attention sur celui-ci), il y a encore un scintillement. C'est seulement vers 75 Hz qu'il disparaît et à 85 Hz l'image est totalement stable. Il est recommandé de régler le taux de rafraîchissement d'un écran à tube cathodique à ces fréquences pour éviter la fatigue des yeux (et de la tête). Le problème du scintillement ne se pose pas avec les écrans LCD. En effet, des tubes fluorescents ou des diodes électroluminescentes produisant une lumière continue éclairent ces écrans par l'arrière. Quelle que soit la fréquence de rafraichissement des données affichées à l'écran, aucun scintillement n'est perceptible[réf. souhaitée].
- Effet phi qui a lieu même avec une succession peu rapide d'images (10 images par seconde). Si l'on dessine une animation sur un carnet et qu'on feuillette les pages, on peut obtenir une illusion de mouvement. Par exemple, les dessins animés ont parfois peu d'images par seconde.
Apprentissage de la vision
La vision est associée à des processus psychologiques très complexes. Dès 1937, Piaget indique qu'elle est chez l'enfant une mise à jour visuelle permanente de la représentation mentale du monde réel de l'individu[6].
Voir aussi
Bibliographie
- Richard Langton Gregory, L'œil et le cerveau : la psychologie de la vision [« Eye and Brain: The Psychology of Seeing »], De Boeck Université, (1re éd. 1966).
- Richard Gregory, Eye and Brain : The psychology of seeing, Princeton University Press, , 5e éd..
- (en) Richard Gregory, Seeing through illusions, Oxford University Press, .
- Hermann von Helmholtz, Optique physiologique, Paris, Masson, (lire en ligne)
- Hermann von Helmholtz, Handbuch der physiologischen Optik, Leopold Voss, (lire en ligne)
Articles connexes
- Vue
- Champ de vision
- Balayage visuel
- Perspective (perception visuelle), Perspective conique
- Illusion d'optique
- Lunettes de vue
- Vision par ordinateur, Vision industrielle
Liens externes
- Conférence expérimentale sur la vision de l'espace Pierre-Gilles de Gennes de l'ESPCI ParisTech
Notes et références
Notes
- ↑ La grandeur caractéristique des rayonnements est leur fréquence, une expression de l'énergie des photons. La longueur d'onde effective varie, avec la vitesse de la lumière, selon le milieu. Par commodité expérimentale, on s'est habitué à indiquer la longueur d'onde dans le vide correspondante.
- ↑ Les lettres désignent, en anglais, le groupe de longueurs d'ondes auxquelles le cône est sensible ; « L : long ; M : medium ; S : short », soit en français L pour longue (faible énergie), M pour moyennes et S pour serré (littéralement, court).
Références bibliographiques
- ↑ Gerald Jacobs et Jeremy Nathans, « L'évolution de la vision des couleurs chez les primates », Pour la Science, no 389, (lire en ligne)
- ↑ (Jordan & Mollon, 1993) http://www.bios.niu.edu/hahin/bios481/tovee3.pdf
- ↑ •(en) Kimberley A. Jameson, « Tetrachromatic color vision », sur aris.ss.uci.edu ;
• Backhaus, Kliegl & Werner « Color vision, perspectives from different disciplines » (De Gruyter, 1998), p. 115-116, section 5.5. ;
• Pr Mollon (université de Cambridge), Pr Jordan (université de Newcastle) « Study of women heterozygote for colour difficiency » (Vision Research, 1993) - ↑ Jean-Baptiste Durand, Yves Trotter, Simona Celebrini ; Privileged processing of the straight-ahead direction in primate area V1, Revue Neuron, 2010/O4/14 (Résumé)
- ↑ Gregory 2000 Ch. 6 « Voir le mouvement ».
- ↑ Jean Piaget, La construction du réel chez l'enfant, Neuchâtel; Paris, Delachaux et Niestlé, (lire en ligne). (Autres éditions au contenu identique, chez le même éditeur: 2e éd. 1950, 3e éd. 1963, 4e éd. 1967, 5e éd. 1973, 6e éd. 1977, 1991).
 Portail de la biologie
Portail de la biologie  Portail de l’œil et de la vue
Portail de l’œil et de la vue  Portail des couleurs
Portail des couleurs