{kind=link}
Eubacteria
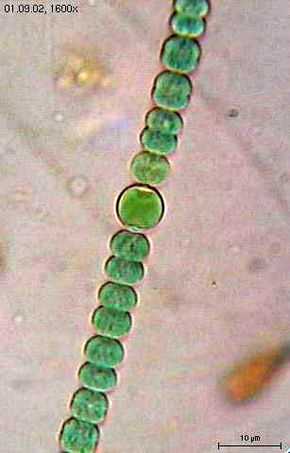
Anabaena sperica, une cyanobactérie
Domaine
Eubacteria
Woese & Fox, 1977
Position :
- Eubactéries
- Protéobactéries
- groupe :
- Firmicutes
- groupe Bacillus / Clostridium
- Actinobactéries
- groupe :
- Cyanobactéries
- groupe Thermus / Deinococcus
- Firmicutes
- groupe :
- Bactéries vertes sulfureuses
- groupe :
- Flavobactéries
- Bactéroïdes
- groupe :
- Spirochètes
- Chlamydiales
- Planctomycétales
- Bactéries vertes non sulfureuses
- Aquificales
- Thermotogales
Les eubactéries (ou Eubacteria), ou « bactéries vraies », sont un groupe majeur de procaryotes. Selon la théorie des trois domaines[1], les eubactéries forment un des trois grands clades qui divisent le monde vivant, les deux autres étant les archébactéries (un autre groupe de procaryotes) et les eucaryotes. Pour d'autres spécialistes de la systématique, les eubactéries sont beaucoup plus vieilles que les deux autres domaines, elle ne constitueraient donc pas un groupe holophylétique mais un grade primitif dans l'histoire de la vie[2],[3],[4].
On peut citer par exemple les cyanobactéries (desquelles proviennent les chloroplastes selon la théorie endosymbiotique), les protéobactéries (desquelles dérivent les mitochondries selon la même théorie), les mycoplasmes (Mollicutes), les entérobactéries, les pseudomonades et la plupart des bactéries à Gram positif.
Étymologie
Création du taxon en 1977
Renommage en 1990 non accepté par tous les biologistes.
Particularités
Les eubactéries se distinguent d’une part des eucaryotes par leur structure cellulaire (la structure procaryote) dont une des caractéristiques est d'être sans noyau, et d’autre part des archées par divers aspects chimiques (dont la structure de la membrane cellulaire) et génétiques.
Leur taille varie de celle d'un virus, comme certaines nanobactéries (0,05 µm) à une taille supérieure à celle de la moyenne des Protozoaires (Epulopiscium fishelsoni (600 µm x 80 µm) est beaucoup plus grande qu'une Paramécie)[5].
Mobilité


Beaucoup de bactéries possèdent des structures extracellulaires comme :
- des flagelles utilisés pour la mobilité des cellules. Les bactéries hétérotrophes peuvent utiliser leurs flagelles pour se diriger vers des zones riches en substances organiques (nutriments) grâce au phénomène appelé chimiotactisme.
- des fimbriae permettant l’attachement
- des pili sexuels indispensables au phénomène de conjugaison.
Quelques bactéries peuvent fabriquer de fines couches externes à la paroi cellulaire, généralement essentiellement constituées de polysaccharides (des sucres).
- Quand la couche est compacte et propre à chaque bactérie, on parle de capsule. Les capsules constituent par exemple une barrière de protection de la cellule contre l’environnement externe et aussi contre la phagocytose. Elle facilite parfois l'attachement aux surfaces (et donc la formation de biofilms). Klebsiella, Bacillus anthracis, Streptococcus pneumoniae sont des exemples de bactéries capsulées.
- Quand la couche est diffuse, on parle de couche mucoïde.
- Quand la couche est plus épaisse, on parle de glycocalyx. Le glycocalyx permet aussi aux bactéries d’adhérer à un support.
- quand la couche forme un tube dans lequel les bactéries sont protégées, on parle de « bactéries engainées ». La gaine est une couche externe dense, transparente et relativement rigide. Ce phénomène est courant chez quelques bactéries aquatiques qui forment des chaînes filamenteuses (Sphaerotilus natans typiquement). La gaine protège les cellules des turbulences de l’eau et permet une vie coloniale.
Les bactéries du groupe Cytophaga – Flavobacterium produisent une couche muqueuse qui leur permet de rester en contact étroit avec un milieu solide. D'autres bactéries comme les Spirillum peuvent s’envelopper d’une couche protéique appelée la couche S.

Certaines bactéries sont mobiles et peuvent se déplacer grâce à des flagelles. Les flagelles des bactéries sont de longs appendices protéiques flexibles. Leur nombre et leur position peuvent différer selon les espèces. La flagellation (ou ciliature) polaire monotriche correspond à la présence d’un seul flagelle à un pôle de la bactérie (exemple des Vibrio). La flagellation polaire lophotriche correspond à la présence de plusieurs flagelles au pôle de la bactérie (Pseudomonas). D’autres bactéries comme Escherichia coli produisent des flagelles sur toute la surface cellulaire et possèdent donc une flagellation péritriche.
Beaucoup de bactéries (comme Escherichia coli) ont deux modes distincts de circulation : mouvement vers l'avant (natation) et mouvement de rotation ou mouvement de « roulis » également et appelé « lang » en anglais. Le tumbling leur permet de se réorienter et leur fait faire un mouvement en trois dimensions, évoluant vers un mouvement de marche aléatoire[6]. Les flagelles d'un groupe de bactéries, les spirochaetes, se trouvent entre deux membranes dans l'espace périplasmique[7].
Le filament du flagelle est constitué d’une protéine, la flagelline. Le type de rotation du flagelle peut déterminer le type de mouvement de la bactérie.
Les bactéries mobiles peuvent réagir à des stimuli, être attirées par des substances nutritives comme les sucres, les acides aminés, l’oxygène, ou être repoussées par des substances nuisibles. Il s'agit de la chimiotaxie, phototaxie et magnétotaxie[8],[9]. Ce comportement est appelé le chimiotactisme. Des chimiorécepteurs de nature protéique sont présents au niveau de la membrane plasmique et du périplasme des bactéries et peuvent détecter différentes substances attractives ou nocives.
Les ions de cuivre bloquent la rotation des flagelles. Pour le faire repartir, on recourt à l’acide éthylènediaminetétraacétique, capable de capturer les ions et donc d'en libérer le flagelle.
Classification
La classification traditionnelle distingue, sur la base du métabolisme, les bactéries photosynthétiques et les bactéries chimiotrophes.
La coloration de Gram est un facteur déterminant dans la taxinomie (classification classique) bactérienne. Elle permet une discrimination entre eubactéries à Gram positif et à Gram négatif.
La systématique moderne, basée sur les analyses génétiques, distingue de nombreux groupes. On notera, entre autres, les cyanobactéries, un groupe important de bactéries photosynthétiques.
Signification évolutive
Selon Thomas Cavalier-Smith, elles forment un groupe paraphylétique (les Neomura, regroupant Archées et Eucaryotes, s'origineraient en leur sein), alors que, classiquement depuis Carl Woese, elles constituent seules l'un des trois empires du vivant. La classification de Woese est privilégiée par les microbiologistes sur base des analyses phylogénétiques. Lors de la découverte des Archées, leur capacité à se développer dans des environnements classiquement défavorables à la vie laissait supposer que les Archées étaient apparues avant les Bactéries, ce qui justifiait la dénomination d'Archéobactéries (vieilles bactéries). Les Bactéries avaient été alors renommées Eubactéries (vraies bactéries). Entre temps, les analyses phylogénétiques ont démontré que les Archées sont phylogénétiquement plus proches des Eucaryotes que des Bactéries et qu'elles étaient apparues après les Bactéries.
En tant que domaine
Modèle à trois domaines proposée par Carl Woese, paradigme majoritairement admis.
En tant que grade
Théorie alternative de Thomas Cavalier-Smith, Gupta, Valas et Bourne
Notion de grade évolutif.
Habitat et écologie

Historiquement, et peut-être encore dans l'esprit général, les bactéries sont associées aux maladies. En effet, de nombreux parasites sont des bactéries (l'inverse n'est pas vrai). L'étude des bactéries a longtemps été dominée par la pathologie, et, dans une moindre mesure, les applications techniques (entre autres la fermentation, que ce soit pour la favoriser ou la combattre).
On trouve des eubactéries dans presque tous les milieux où la vie se révèle possible. Leur répartition va de l'atmosphère aux limites inférieures des sols et des fonds océaniques en passant par tous les écosystèmes.
Selon les lignées, on en trouve des phototrophes (tirant leur énergie de la lumière), chimiotrophes (trouvant leur énergie de gradients chimiques non organiques), hétérotrophes (trouvant leur énergie dans la matière organique, qu'elle soit vivante (parasitisme) ou morte (saprophytisme)); on en trouve des aérobies ou des anaérobies, strictes ou non.
Notes et références
- ↑ Carl Richard Woese & George Edward Fox, 1977. Phylogenetic structure of the prokaryotic domain: the primary kingdoms. Proc Natl Acad Sci USA. 74(11):5088–5090
- ↑ Thomas Cavalier-Smith, 2006. Cell evolution and Earth history: stasis and revolution. Philos Trans R Soc Lond B Biol Sci, 361(1470):969-1006.
- ↑ Thomas Cavalier-Smith, 2010. Deep phylogeny, ancestral groups and the four ages of life. Philos Trans R Soc Lond B Biol Sci, 365(1537):111-32.
- ↑ Valas RE & Bourne PE, 2011. The origin of a derived superkingdom: how a gram-positive bacterium crossed the desert to become an archaeon. Biol Direct 6:16
- ↑ Lecointre G. Le Guyader H. Classification phylogénétique du vivant, Belin 2001 ISBN 2-7011-2137-X
- ↑ Wu M, Roberts J, Kim S, Koch D, DeLisa M, « Collective bacterial dynamics revealed using a three-dimensional population-scale defocused particle tracking technique », Appl Environ Microbiol, vol. 72, no 7, , p. 4987–94 (PMID 16820497, DOI 10.1128/AEM.00158-06, lire en ligne)
- ↑ Bardy S, Ng S, Jarrell K, « Prokaryotic motility structures », Microbiology, vol. 149, no Pt 2, , p. 295–304 (PMID 12624192, DOI 10.1099/mic.0.25948-0, lire en ligne)
- ↑ Lux R, Shi W, « Chemotaxis-guided movements in bacteria », Crit Rev Oral Biol Med, vol. 15, no 4, , p. 207–20 (PMID 15284186)
- ↑ Frankel R, Bazylinski D, Johnson M, Taylor B, « Magneto-aerotaxis in marine coccoid bacteria », Biophys J, vol. 73, no 2, , p. 994–1000 (PMID 9251816)
Liens externes
- Référence Tree of Life Web Project : Eubacteria (en) (consulté le 30 juil. 2012)
- Référence Fossilworks Paleobiology Database : Eubacteria (en) (consulté le 30 juil. 2012)
Voir aussi
 Portail de la microbiologie
Portail de la microbiologie  Portail de l’évolution
Portail de l’évolution