
{kind=link}
Cladistique
|
|
Des informations de cet article ou section devraient être mieux reliées aux sources mentionnées dans la bibliographie ou en liens externes. (février 2013) Améliorez sa vérifiabilité en les associant par des références.
|
La cladistique (ou le cladisme[1], du grec ancien κλάδος, klados, signifiant « branche »), aussi appelée systématique phylogénétique, est une théorie de classification phylogénétique.
Elle classe les êtres vivants selon leurs relations de parenté, dans un cadre évolutionniste. Elle repose sur la construction de groupes monophylétiques dits clades qui incluent un ancêtre commun et l'ensemble de sa descendance.

Corpus théorique

La cladistique cherche à établir les relations de parenté entre les taxons, sur la base d'un partage d'états de caractères considérés pertinents (états dits apomorphes, par opposition aux états plésiomorphes qui sont des états partagés mais supposés non-pertinents pour regrouper les taxons). La cladistique fournit le cadre théorique du test à faire passer aux « états » afin de rejeter ou non leur pertinence. Un état non-rejeté est appelé synapomorphe et sert à caractériser un groupe monophylétique.
Cette théorie a été présentée dans les années 1950 par l'entomologiste allemand Willi Hennig.
Le résultat de la reconstruction phylogénétique est représenté sous forme d'un graphe connexe non cyclique, ou arbre, appelé ici cladogramme. Un tel arbre indique les relations de parenté entre les taxons considérés. Des connaissances morphologiques, moléculaires et comportementales sont utilisées pour les travaux cladistiques, largement assistés par des programmes informatiques.
Concepts importants de la cladistique
Le clade comme unité de classification
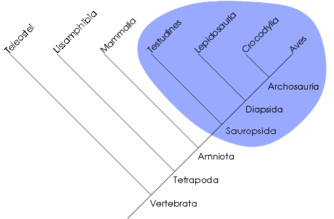
La cladistique reconnaît une unité de classement bien particulière, le clade. Un clade est un ensemble d'organismes formant un groupe monophylétique, c'est-à-dire une totalité de descendance (un ancêtre commun et tous ses descendants). Ce concept ne s'oppose pas à celui de grade, rapprochement d'organismes reposant sur d'autres critères (par ex. ressemblance générale, somme de modifications adaptatives). Cependant, la cladistique qualifie les grades de groupes paraphylétiques ou polyphylétiques selon que le rapprochement est effectué sur la base de plésiomorphies ou d'homoplasies. Certains grades peuvent toutefois être monophylétiques. Par exemple, les algues forment un grade polyphylétique, les reptiles forment un grade paraphylétique et les mammifères forment un grade holophylétique.
Un clade peut aussi être défini comme un ensemble d'organismes phylogénétiquement plus proches les uns des autres qu'ils ne le sont d'aucun autre organisme. Par exemple, les reptiles ne forment pas un clade car certains (les crocodiliens) sont plus apparentés aux oiseaux (formant ainsi le clade archosauriens) qu'aux autres reptiles. Ainsi, un clade correspond à une unité évolutive. On parle aussi parfois de groupe holophylétique, ce qui est synonyme de groupe monophylétique sensu Hennig.
Par définition le taxon est l'unité des classifications scientifiques du vivant. Dans le cadre de la théorie cladistique, tous les taxons sont des clades et tous les clades sont des taxons, contrairement à la taxonomie numérique ou à la systématique évolutionniste où un grade non holophylétique peut être un taxon.
Il est parfois fait mention d'« espèce ancestrale » pour désigner une espèce représentante d'un clade. Cependant, une telle espèce, si elle existait, serait nécessairement définie sur la base d'états de caractères plésiomorphes par rapport à ses descendants. Elle correspondrait alors à la définition d'un groupe paraphylétique et ne serait pas reconnue comme unité évolutive par la théorie cladistique. Dans ce cadre théorique, l'ancêtre est, premièrement, hypothétique et, deuxièmement, n'est pas caractérisable en tant que taxon. « Non caractérisable » ne signifie pas « non identifiable » : il est donc possible de dresser un morphotype ancestral.
Paraphylie

La notion de paraphylie s'applique à un groupe défini sur le partage d'un ou de plusieurs états de caractères plésiomorphes. Les taxons de ce genre de groupes ne constituent pas une totalité de descendance puisque les organismes portant les états apomorphes en sont exclus. Un groupe paraphylétique n'est donc pas monophylétique. La théorie cladistique ne reconnaît pas la pertinence de groupes tel que les reptiles, les poissons, les invertébrés, les procaryotes, etc. Au sein d'un groupe paraphylétique, les taxons ne sont pas forcément plus apparentés entre eux qu'à d'autres taxons. Par exemple, les tétrapodes sont un groupe monophylétique au sein d'un groupe plus général comprenant aussi les poissons. Certains poissons comme le cœlacanthe sont plus proches des tétrapodes qu'ils ne le sont d'autres poissons tels le requin. Ainsi, le terme poisson désigne un groupe paraphylétique.
Le cladogramme étant le résultat d'un test de monophylie, c'est lui qui indique si un groupe est monophylétique ou paraphylétique. Il est donc important de justifier le choix des états de caractères considérés, en amont de l'analyse, apomorphes ou plésiomorphes (le test de monophylie ne concernant que les états apomorphes). Pour cela, il existe plusieurs critères dont celui du groupe externe (le plus utilisé), le critère ontogénétique et à moindre mesure les critères paléontologique et chorologique. Soient deux états a et b d'un même caractère dont la relation peut être schématisée par a↔b en l'absence de justification sur l'apormophie ou la plésiomorphie de l'un ou de l'autre. Le schéma a→b indiquerait a comme étant plésiomorphe et b apomorphe. Le schéma b→a indiquerait le contraire.
Homologie versus homoplasie

On distingue deux types de ressemblances, l'homologie et l'homoplasie. Selon la définition la plus courante, des états de caractères dits homologues sont hérités d'un ancêtre commun. Pour quelques auteurs, la relation entre états est appelée homologie. Deux états homologues sont non seulement ressemblants, mais surtout phylogénétiquement « identiques ». À l'inverse, l'homoplasie, terme introduit par Lankester en 1870, désigne des états de caractères non hérités d'un ancêtre commun. Des états dits homoplastiques sont donc ressemblants mais ne sont pas phylogénétiquement « identiques ». Savoir distinguer la source de ces ressemblances est une tâche compliquée. Plusieurs critères permettent de proposer le statut « homologues » pour ces ressemblances, le plus utilisé étant le « principe des connexions » d'Étienne Geoffroy Saint-Hilaire (ensuite repris par Richard Owen) appelé aussi « identité des connexions » : deux organes sont homologues si, quelles que soient leurs formes et/ou fonctions, ils ont les mêmes connexions avec d'autres organes. Ce principe peut aisément s'étendre aux séquences moléculaires et sous-tend la pratique d'alignement des séquences.
Ce principe ne permet que de formuler des hypothèses dites d'homologie primaire. En effet rien ne nous assure que les états de caractère supposés homologues le sont effectivement. La cladistique offre le cadre théorique pour tester de telles hypothèses. Il s'agit d'un test de congruence selon Hennig ou de parcimonie. Le principe de congruence (ou de parcimonie) cherche à maximiser le nombre d'hypothèses d'homologie compatibles entre elles afin de minimiser le nombre d'hypothèses surnuméraires (ou ad hoc), une hypothèse ad hoc étant une hypothèse d'homoplasie (pouvant recevoir quantité d'explications la rendant non testable). Le cladogramme reconstruit est le résultat de la maximisation de la congruence parmi les hypothèses d'homologie. Une hypothèse non rejetée par le test est appelée « homologie secondaire ».
À l'inverse, une hypothèse d'homologie rejetée par le test permet d'attribuer le statut d'homoplasie aux états en question. N'importe quelle explication permettrait de comprendre l'origine des homoplasies. Deux explications principales sont souvent discutées : la convergence évolutive et la réversion. La convergence indique qu'une ressemblance est apparue plusieurs fois indépendamment. La réversion est interprétée comme étant la perte secondaire d'un état de caractère, c'est-à-dire le retour à un état ressemblant à l'état plésiomorphe. Par exemple tous les Vertébrés ne possèdent pas de membres pairs. Or l'ensemble des Vertébrés concernés par cette absence ne forme pas un groupe monophylétique. On dit par exemple des Gymnophiones ou des Serpents qu'ils ont « perdu » ces membres pairs. L'homoplasie n'étant pas un caractère hérité par un ancêtre commun, elle ne nous renseigne pas sur les relations de parenté. Un groupe identifié sur la base d'une homoplasie est appelé groupe polyphylétique.
Dans certains cas il n'y a aucun élément pour savoir si l'homoplasie est une réversion ou une convergence, on parle alors d'interprétation ambiguë[2]. Le choix réversion ou convergence est arbitraire[3]. L'hypothèse ACCTRAN (Accelerated transformation) favorise les réversions. L'hypothèse DELTRAN (Delayed transformation) favorise les convergences. L'image ci-dessous l'illustre pour deux arbres ayant la même répartition des caractères.
À titre d'exemple, l'absence de fenêtre antéorbitaire chez les crocodiles actuels est considéré comme une réversion (elle était présente chez leurs ancêtres putatifs) ; les topologies similaires du bassin des oiseaux et des ornitischiens, ou encore les formes hydrodynamiques des delphinidés et de la plupart des lamniformes sont considérées comme des convergences.

Lecture d'un cladogramme
L'attribution des états de caractères aux taxons est représentée, le plus généralement, dans un tableau appelé matrice taxons-caractères. Voici la matrice hypothétique dans laquelle, pour chaque caractère "x", l'état plésiomorphe est noté x et l'état apomorphe x.
| caractère | taxon A | taxon B | taxon C | taxon D | taxon E | taxon F |
|---|---|---|---|---|---|---|
| Caractère "a" | a | a | a' | a' | a' | a' |
| Caractère "b" | b | b | b' | b | b | b |
| Caractère "c" | c' | c | c | c | c' | c' |
| Caractère "d" | d | d' | d' | d' | d | d' |
| Caractère "e" | e | e | e | e | e | e |
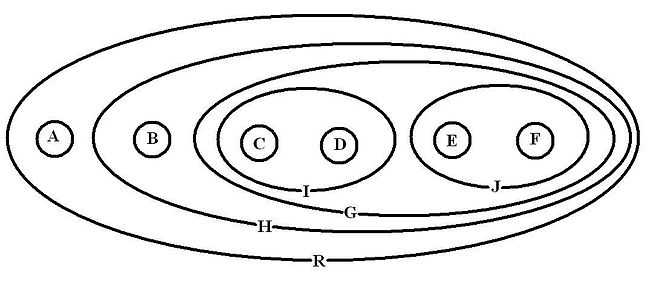
Soient les taxons A, B, C, D, E et F. On considère l'arbre suivant: (A(B((C, D)(E, F)))). Ici R={A+B+C+D+E+F} : c'est la racine ou le nœud qui contient tout. H= {B+C+D+E+F} G= {C+D+E+F} = {I, J} avec I= {C+D} et J= {E+F}. Les nœuds internes H, G, I et J sont des taxons au même titre qu'A, B, C, D, E et F.
L'histoire parcimonieuse des états de caractères est représentée par des barres rouges où x→x' indique le passage de l'état x à x' et x'→x le passage de l'état x' à l'état x. Le passage d'un état à un autre est appelé transformation ou pas évolutif.

Ici, l'état a' est commun aux taxons C, D, E, et F; c'est donc une synapomorphie de G.
L'état b' n'apparaît que sur le seul taxon terminal C. C'est donc une autapomorphie de C. Cet état ne renseigne pas sur les relations de parenté de C avec les autres taxons.
L'état e est commun à tous les taxons, c'est donc une symplésiomorphie.
L'état c' apparaît deux fois dans l'arbre. Le passage de c à c' coûte donc deux pas évolutifs. L'état c' est une homoplasie (il n'est donc pas interprété comme hérité d'un ancêtre commun).
Le passage de l'état d à d' est suivi d'un autre passage inverse de d' à d. Cette deuxième transformation est généralement interprétée comme une réversion.
Le coût total des transformations est de 6 pas évolutifs.
On dit de deux taxons qu'ils sont groupes-frères quand ils sont plus proches entre eux que de n'importe quel troisième taxon. Ici par exemple, C et D sont groupes-frères, ainsi que B et G ou encore J et I.
Un cladogramme est un graphe particulier, appelé hiérarchie en mathématiques. Une hiérarchie est équivalente à un emboîtement de classes. Chaque classe correspond à un nœud du graphe. Au sens phylogénétique, un nœud ou une classe est un taxon. Ici les taxons C et D sont inclus dans une classe I. Tous les taxons (terminaux ou inclusifs) sont inclus dans R : la définition de la racine est d'être la classe incluant toutes les autres classes.
Un graphe hiérarchique peut être représenté par un diagramme de Venn pour faire apparaître l'emboîtement des classes. L'image ci-dessous représente le diagramme de Venn correspondant au cladogramme utilisé dans l'exemple précédent.
Enracinement et groupe externe
Le principe dit du « groupe externe » ou « extra-groupe » est généralement utilisé pour enraciner un arbre phylogénétique, c'est-à-dire désigner le nœud correspondant à la classe racine (contenant toutes les autres classes)[4]. On estime que tout état de caractère observé en dehors du groupe interne est plésiomorphe pour le groupe interne. Au contraire, tout état propre à des taxons du groupe d'étude est considéré apomorphe.
Une des méthodes mettant en œuvre ce principe introduit un taxon du groupe externe dans l'analyse : par exemple un téléostéen pour une étude phylogénétique des tétrapodes ; un céphalopode ou un bivalve pour une étude phylogénétique des gastéropodes. Il faut veiller à ce que les caractères des groupes externe et interne soient comparables, ce qui incite à choisir un groupe externe relativement proche du groupe d'étude. Dans l'image ci-dessous, l'arbre de départ n'est pas enraciné alors que les deux arbres ci-dessous (sous forme de graphe ou de diagramme de Venn) sont enracinés, c'est-à-dire orientés grâce aux groupes externes A ou C.
Les relations de parenté et le sens des transformations de caractères ne peuvent être inférées qu'à partir d'un arbre enraciné (autrement dit : du cladogramme).
On remarque que le choix du groupe externe (ici A ou C) modifie la topologie de l'arbre.
Il arrive que la méthode décrite ci-dessus outrepasse le principe énoncé. En effet, le taxon du groupe externe introduit dans l'analyse ne présente pas nécessairement la totalité des caractères sous l'état plésiomorphe.
D'autres méthodes d'enracinement proposent d'appliquer ce principe pour chaque caractère, en comparant le groupe d'étude à plusieurs taxons du groupe externe. Ces taxons ne sont alors pas introduits dans l'analyse :
- soit un taxon hypothétique est reconstruit ;
- soit les caractères sont polarisés en amont de l'analyse (dans le cas d'une représentation par matrice) ;
- soit les caractères sont hiérarchisés en amont de l'analyse (dans le cas d'une représentation par hiérarchie).

Cladistique et évolution

Deux notions ont été proposées pour distinguer les écoles de pensée au sein de la cladistique : l'école pattern et l'école process. Le pattern fournit la structure représentant les relations de parenté (c'est-à-dire le graphe hiérarchique ou le diagramme de Venn). Le process fait appel aux processus de évolution (ex. : sélection naturelle, spéciation, adaptation…).
Les cladogrammes résultant d'analyses cladistes fournissent les clades et le statut des états de caractères. La théorie de l'évolution permet d'interpréter leur évolution.
Méthodes cladistiques
Différentes méthodes de reconstruction phylogénétique existent en cladistique. Le principe de parcimonie, selon lequel l'explication la plus économe en hypothèses doit être retenue, les sous-tend. Ce principe est attribué à Guillaume d'Ockham (les entités ne doivent pas être multipliées par delà ce qui est nécessaire : « Entia non sunt multiplicanda praeter necessitatem »).
Ces méthodes sont :
- la parcimonie (dite standard),
- l'optimisation directe,
- l'analyse à trois éléments (3ia)[5]
- la compatibilité (aujourd'hui tombée en désuétude).
Hennig, le fondateur de la théorie cladistique, n'a jamais utilisé ces méthodes aujourd'hui assistée par ordinateur. Il parlait de méthode d'argumentation. Aujourd'hui une phylogénie n'est que très rarement acceptée sans analyse algorithmique pour l'appuyer.
Parcimonie
Selon la méthode de parcimonie standard, l'économie d'hypothèse touche au nombre de pas évolutifs. L'arbre le plus court (c'est-à-dire l'arbre avec le moins de pas évolutifs) est l'arbre représentant l'hypothèse phylogénétique la plus acceptable. Le comptage des pas peut-être influencé par la considération que l'on a des convergences et/ou réversions, d'où l'existence de plusieurs écoles méthodologiques :
- La parcimonie de Wagner (convergences et réversions sont acceptées).
- La parcimonie de Camin-Sokal (les convergences sont admises mais pas les réversions).
- La parcimonie de Dollo (les réversions sont admises mais pas les convergences).
Optimisation directe
Selon la méthode d'optimisation directe, l'économie d'hypothèses se situe en amont de l'analyse, dans le codage même des caractères. Techniquement, proposer les caractères conduisant à l'arbre parcimonieux s'effectue en même temps que la recherche de cet arbre (c'est-à-dire lors du comptage des pas). Dans un cadre d'analyse moléculaire, cette méthode ne nécessite pas d'alignement des séquences préalablement à l'analyse. Les nucléotides de séquences alignées sont équivalents au codage d'états de caractères identiques. Ici, l'alignement (ou le codage) est donc effectué au fur et à mesure de l'analyse de manière à optimiser le nombre de pas. L'arbre obtenu par optimisation directe est théoriquement plus court qu'un arbre obtenu par une autre méthode d'alignement .
Compatibilité
Selon la méthode de compatibilité, l'économie d'hypothèses concerne le nombre de caractères permettant d'éviter les homoplasies. On dit de tels caractères qu'ils sont mutuellement compatibles. L'ensemble des caractères mutuellement compatibles formera une clique. L'arbre retenu sera construit à partir de la clique la plus importante. Il sera donc dépourvu d'homoplasie.
Analyse à trois éléments
L'analyse à trois éléments décompose les caractères en hypothèses relationnelles à trois éléments spécifiant que deux taxons sont plus proches entre eux que d'un troisième. Ces hypothèses minimales sont appelées 3is pour three item statements ou « assertions à trois éléments », d'où le nom de la méthode. Cela implique d'avoir accès aux hypothèses relationnelles (ou de parenté) sur les caractères : les caractères sont donc logiquement représentés sous la forme d'arbres racinés, ou autrement dit, de graphes hiérarchiques. Un caractère consiste à attribuer les états d'une structure morpho-anatomique aux taxons de l'étude après avoir indiqué la relation (hiérarchique) entre ces états. L'arbre phylogénétique reconstruit l'est à partir de l'ensemble le plus grand de 3is compatibles entre eux.
La 3ia a d'abord été utilisée pour la biogéographie historique et a ensuite été utilisée pour l'étude des taxons dans le cadre théorique de la cladistique. Cette méthode n'utilise pas, pour l'analyse, de taxon du groupe externe. Le systématicien applique en amont de l'analyse le principe extra-groupe, entre autres, pour désigner les états informatifs des caractères, c'est-à-dire les états permettant de regrouper des taxons.
Très peu d'études ont comparé la performance le l'analyse à trois éléments à celle de la parcimonie. La plus récente[6] a trouvé que la 3ia donnait une excellente puissance et un taux d'erreurs (clades artéfactuels) intermédiaires entre celui de la parcimonie avec états ordonnés (donnant le moins d'erreurs) et celui de la parcimonie non-ordonnée (donnant le plus d'erreurs).
Argumentation hennigienne
Cette procédure non automatisée consiste à proposer une phylogénie (ou schéma d'argumentation) sur la base d'arguments pour chaque clade, les arguments étant les états de caractères explicitement donnés par l'auteur.
Notes et références
- ↑ http://www.larousse.fr/encyclopedie/nom-commun-nom/cladistique/34254
- ↑ M.G.G. de Pinna, Concepts and tests of homology in the cladistic paradigm, Cladistics, 1991.En ligne (en)
- ↑ I. Agnarson & J. A. Miller Is ACCTRAN better than DELTRAN?, Cladistics, 2008 En ligne (en)
- ↑ V. Barriel & P. Tassy Rooting with Multiple Outgroup: Consensus Versus Parsimony, Cladistics, 1998
- ↑ R. Zaragüeta-Bagils & E. Bourdon, Three-item analysis: Hierarchical representation and treatment of missing and inapplicable data, Comptes Rendus Palevol, Volume 6, Issues 6-7, November 2007, Pages 527-534
- ↑ Grand A, Corvez A, Duque Velez LM, Laurin M. (2013). Phylogenetic inference using discrete characters: performance of ordered and unordered parsimony and of three-item statements. Biological Journal of the Linnean Society, 110 (4): 914–930.
Voir aussi
Bibliographie
- P. Tassy, La renaissance de la systématique, 2001 En ligne (fr)
- H. Le Guyader, G. Lecointre, Classification phylogénétique du vivant 3ème édition, Belin, 2001.
- P. Tassy L'arbre à remonter le temps, Christian Bourgois, Paris, 1991.
- P. Darlu & P. Tassy, La reconstruction phylogénétique. Concepts et Méthodes, Masson, 1993 En ligne (fr)
- W. Hennig, Phylogenetic Systematics, Illinois University Press, 1966, traduit par D. Dwight Davis & R. Zangerl.
- I.J. Kitching, P.L. Forey, C.J. Humphries et D.M. Williams, Cladistics, Oxford, Oxford University Press, 1998 (2e éd.).
- C. Patterson, Morphological characters and homology, in K.A. Joysey et A.E. Friday (éd.), Problems in Phylogenetic Reconstruction, Londres, Academic Press, 1982.
- K. de Queiroz et J.A. Gauthier, « Phylogenetic taxonomy », Annual Review of Ecology and Systematics no 23, 1992, p. 449–480.
- D.L. Swofford, G.J. Olsen, P.J. Waddell et D.M. Hillis, Phylogenetic inference, in D.M. Hillis, C. Moritz et B.K. Mable (éd.), Molecular Systematics, Sunderland (Massachusetts), Sinauer Associates, 1996.
- E.O. Wiley, Phylogenetics : The Theory and Practice of Phylogenetic Systematics.-, New York, Wiley Interscience, 1981.
Articles connexes
- Caractère ancestral et caractère dérivé
- Willi Hennig
- Monophylie
- Paraphylie
- Polyphylie
- Synapomorphie
- Autapomorphie
- Symplésiomorphie
- Analyse à trois éléments
Lien externe
- (en) The Willi Hennig Society website
 Portail de la biologie
Portail de la biologie  Portail de l’évolution
Portail de l’évolution