
{kind=link}
Théorie de la récapitulation
La récapitulation est une théorie en biologie de l’évolution du développement développée par Ernst Haeckel qui veut que l’ontogénie d’un organisme passe par des stades représentant les espèces ancestrales de celui-ci. Haeckel a résumé la théorie par la phrase « l’ontogenèse récapitule la phylogenèse ».
Origines
La théorie du parallélisme
La théorie du parallélisme établie par Johann Meckel et Étienne Serres démontrait qu’un animal avait comme phase développementale la phase adulte de l’espèce inférieure[1]. Les espèces inférieures seraient donc en fait des formes développementales des espèces plus supérieures. Comme cette théorie est apparue avant l’arrivée de l’évolutionnisme de Darwin, les termes «inférieur» et «supérieur» font références à la chaîne des êtres (Great Chain of Being)[2]. Cette théorie fut rapidement rejetée par plusieurs biologistes dont Karl Ernst von Baer.
La divergence de von Baer
Karl Ernst von Baer, considéré comme le père de l’embryologie, a publié plusieurs ouvrages dont un en 1828 qui stipule que les embryons de chaque organisme ont un stade initial semblable puis divergent peu à peu pour former l’adulte. Il remarqua aussi que les caractères généraux à un certain groupe apparaissent plus tôt dans le développement que les caractères plus spécialisés. Une de ses lois explique que l’embryon d’un animal ne ressemble jamais au stade adulte d’un autre animal même s’il est moins évolué. L’embryon ressemble seulement à l’autre embryon[3].
La théorie de Haeckel
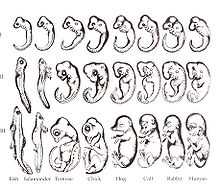
Dans son livre "Generelle Morphologie der Organismen" paru en 1866, Ernst Haeckel développa la théorie de la récapitulation aussi appelée la «loi biogénétique fondamentale»[4]. Il s’est inspiré de la théorie de Meckel-Serres en ajoutant des éléments du Darwinisme pour former sa théorie[3]. Le fondement de sa théorie est que chaque organisme se développe en passant par les étapes des organismes ancestraux. Par exemple, un oiseau se développerait en passant par tous les phénotypes de ses ancêtres en passant par une forme aquatique invertébrée, une forme de type poisson, une forme de type amphibien, etc. Des centaines de millions d’années d’évolution seraient donc résumés dans l’ontogénie des animaux. Avec l’addition terminale, Haeckel démontre que chaque caractère qui apparait chez un organisme s’ajoute à la fin des étapes développementales des descendants[3]. Deux types d’embryogénies furent créées par Heckel : la cénogenèse, pour expliquer l’ontogénie raccourcie ou falsifiée, et la palingenèse, pour l’embryogénie expliquant parfaitement la phylogenèse[5]. Haeckel a aussi réalisé plusieurs croquis d’embryons pour illustrer sa théorie. Plusieurs critiquèrent ses dessins en lui reprochant d’avoir exagéré les traits des embryons et d'avoir dessiné des caractères inventés qui permettaient de prouver sa théorie[6]. En 1997, une étude aurait prouvé que Haeckel aurait bel et bien exagéré des similarités entre embryons et qu’il aurait exclut des embryons qui seraient différents[7].
La discréditation de la théorie
Plusieurs biologistes critiquèrent la théorie de Haeckel pendant le XXe siècle. Gavin de Beer écrivit en 1940 que la récapitulation était fausse et qu’elle n’avait jamais lieu [8]. Par contre, Stephen J. Gould a affirmé que la récapitulation se produit assez souvent, mais qu’il ne considère pas la théorie comme une loi, car elle n’est pas assez fréquente[4]. En effet, en observant l’ontogénie de la plupart des organismes, on se rend compte que les phases développementales ne représentent pas le processus évolutif qui a mené à l’espèce actuelle.
Par exemple, les gastéropodes de type sénestre ont évolué à partir des gastéropodes de type dextre. Cependant, dans l’ontogénie de ceux-ci il n’y a pas de passage entre une forme dextre à une forme sénestre. Le développement est en fait altéré au tout début du développement alors que la division des cellules se dirige vers la gauche au lieu d’aller vers la droite [9]. Un gastéropode ne peut donc pas passer de l’état dextre à l’état senestre pendant son développement.
De plus, il existe plusieurs espèces qui sont phylogénétiquement proches et se ressemblent, mais qui ont un mode développemental complètement différent. C’est le cas de certaines espèces d’oursins En effet, certains ont un mode indirect qui passe par une forme larvaire, tandis que d’autres ont un mode direct qui se développe sans larve[10]. L’espèce ancestrale commune aurait un développement indirect ce qui signifie que l’espèce avec un développement direct ne peut pas récapituler la phylogénie, car il y a une partie développementale entière qui est retirée.

Un autre exemple qui prouve que la récapitulation n’est pas universelle est l’axolotl. Cet animal a évolué à partir d’une salamandre en gardant certains caractères juvéniles de celle-ci. Par conséquent, son développement est sensiblement le même que celle de son ancêtre, mais il se termine de façon précoce. Ce phénomène se nomme la néoténie. Le système reproducteur de l'organisme devient mature avant que le développement somatique atteigne celui de l'ancêtre. Dans ce cas-ci, l'axolotl est mature avec les caractéristiques juvéniles de l'espèce ancestrale comme les branchies externes[11]. L’ontogénie de l’axolotl ne récapitule donc pas toute les étapes de l’évolution menant à lui. En fait, tous les changements dans les caractères qui sont causés par une modification du patron de développement ne peuvent pas présenter une récapitulation, car le nouveau caractère n’est pas ajouté à la fin développement. La récapitulation est donc possible seulement pour les caractères qui s’ajoutent par addition terminale et qui sont causés par des changements mineurs du phénotype[3].


Quelques exemples réels
La récapitulation est considérée maintenant comme un phénomène peu fréquent, mais qui peut se produire chez certains organismes. Ce n’est pas un outil pour étudier l’évolution d’une certaine espèce, car souvent c’est seulement une partie du développement qui est récapitulé et un seul caractère est considéré. Plusieurs exemples démontrent ce phénomène.
Les fentes branchiales des tétrapodes
À un certain moment dans son développement, l’embryon de l’être humain a des fentes branchiales et une queue identiques à celles des poissons. Cette caractéristique est partagée par tous les vertébrés dont le poulet[4]. Ceci permet de constater que les poulets et les humains auraient un ancêtre commun avec les poissons. Cette caractéristique constitue donc une certaine récapitulation de l’évolution de l’espèce malgré le fait que ce ne soit pas une récapitulation précise de tous les changements phénotypiques menant à l’organisme actuel. La récapitulation est donc assez importante, car bien qu’elle soit peu présente elle prouve que des groupes très éloignés ont une même origine[12].
Les membres postérieurs des cétacés
Chez certains cétacés, l’embryon possède à un moment des bourgeons de membres postérieurs. Avec l'évolution, les mammifères marins ont perdu progressivement les pattes postérieures, car ils utilisaient de plus en plus leur queue pour se propulser[13]. L’embryon de ces mammifères récapitule donc leur phylogénie passant d’un mammifère terrestre avec des membres postérieurs à un mammifère marin sans ces membres postérieurs.

Les larves des cirripèdes
Les cirripèdes sont des organismes marins et sessiles qui étaient classés dans les mollusques jusqu’en 1830. Les larves de cirripèdes ont permis de découvrir que ceux-ci sont en fait des crustacés[14]. En effet, malgré leur physionomie au stade adulte très différente des autres crustacés, la larve est similaire à celle des autres crustacés. C’est un exemple de récapitulation, car le stade développemental des cirripèdes démontre que dans le passé ils avaient des caractères de crustacés puis les ont perdus pour bénéficier d’une nouvelle niche écologique. Les larves possèdent des appendices sur le céphalon comme les antennes et les antennules qui sont absents chez les adultes. Le premier stade larvaire des cirripèdes est la larve nauplius qui est un élément caractéristique des crustacés. Cela a donc permis de les classer dans ce groupe[9].
Les yeux des poissons plats
Les poissons plats (Pleuronectiformes) sont aussi un exemple de la récapitulation. En effet, les yeux des pleuronectiformes sont sur le même côté de la tête chez l’adulte. Cette position des yeux sert à pouvoir voir avec leurs deux yeux lorsqu’ils sont à plat sur le fond marin. De plus, les pleuronectiformes maintiennent souvent leur corps à un certain angle au-dessus du substrat pour pouvoir plus facilement piéger leurs proies. Cette position des yeux améliorerait donc l’efficacité de cette technique[15]. Les yeux n’apparaissent pas à cet emplacement lors du développement de l’animal. Lorsque les yeux se forment, ils ont une disposition symétrique comme chez leurs ancêtres. Puis, peu à peu un des deux yeux migre vers la position finale. Ceci est un bon exemple de récapitulation, car l’ontogénie montre le processus qui a permis aux yeux de passer en position symétrique vers une position asymétrique. L’étude de poissons plats plus ancestraux a démontré que ce déplacement s’est bel et bien fait de façon progressive[15].
Le conduit à venin des centipèdes
Selon une étude datant de 2012, les centipèdes du genre Scolopendra seraient aussi un exemple de récapitulation. Effectivement, les conduits à venin se forment chez le centipède immature de la même manière qu’ils apparaissent au cours de l’évolution. Le conduit à venin se développe premièrement comme une rainure à la surface de l’exosquelette des forcipules. Ensuite, la rainure se referme pour former le conduit circulaire attaché à l’endocuticule de l’exosquelette. Finalement, lorsque le centipède devient mature, le conduit se détache de l’exosquelette. C’est le même processus qui s’est produit au fil du temps, prouvé par l’étude de spécimens fossiles[9].
Voir aussi
- Biologie évolutive du développement
- Ontogeny and Phylogeny de l'Université de la Californie Berkeley
- Division de la médecine et de la biologie, Université Brown. "Evolution and Development I: Size and Shape"
Références
- ↑ (en) W. Shumway, « The recapitulation theory », The Quaterly Review of Biology, no 1, , p. 93-99
- ↑ (en) E. Mayr, « Recapitulation reinterpreted: the somatic program », Quarterly Review of Biology, , p. 223–232.
- 1 2 3 4 (en) K.S. Thomson, « Marginalia: Ontogeny and Phylogeny Recapitulated », American Scientist, vol. 76, no 3, , p. 273-275
- 1 2 3 (en) Stephen J. Gould, Ontogeny and Phylogeny, Belknap Press of Harvard University Press,
- ↑ L. Vialleton, « La loi biogénétique de Hackel », Revue de Métaphysique et de Morale, vol. 16, no 4, , p. 448-465
- ↑ (en) M.K. Richardson et G. Keuck, « Haeckel's ABC of evolution and development », Biological Reviews of the Cambridge Philosophical Society, no 4, , p. 495-528
- ↑ (en) M.K. Richardson et al., « There is no highly conserved embryonic stage in the vertebrates: implications for current theories of evolution and development », Anatomy and embryology, vol. 196, no 2,
- ↑ (en) Gavin R. de Beer, Embryos and Ancestors, Oxford, The Clarendon Press,
- 1 2 3 (en) M.M. Dugon, L. Hayden, A. Black et W. Arthur, « Development of the venom ducts in the centipede Scolopendra: an example of recapitulation », Evolution & development, vol. 14, no 6, , p. 515-521
- ↑ (en) « Reproduction and life history », sur l'Echinoid Directory du Natural History Museum (consulté le 20 novembre 2014)
- ↑ (en) « Ambystoma mexicanum », sur Animal Diversity Web (consulté le 20 novembre 2014)
- ↑ Paul Mengal, Histoire du concept de récapitulation: ontogenèse et phylogenèse en biologie et sciences humaines, Masson,
- ↑ (en) J.G.M. Thewissen, M.J. Cohn, L.S. Stevens, S. Bajpai, J. Heyning et W.E. Horton, Jr., « Developmental basis for hind-limb loss in dolphins and origin of the cetacean bodyplan », Proceedings of the National Academy of Sciences, no 22, , p. 8414-8418
- ↑ J. Forest, « Cirripèdes », sur Encyclopædia Universalis (consulté le 12 novembre 2014)
- 1 2 (en) M. Friedman, « The evolutionary origin of flatfish asymmetry », Nature, no 7201, , p. 209-212
 Portail de la biologie
Portail de la biologie  Portail de l’évolution
Portail de l’évolution