Sénescence
|
|
Cet article est une ébauche concernant la médecine et la biologie. Vous pouvez partager vos connaissances en l’améliorant (comment ?) selon les recommandations des projets correspondants.
|
En biologie, la sénescence (du latin : senex, « vieil homme » ou « grand âge »), ou vieillissement, est un processus physiologique qui entraîne une lente dégradation des fonctions de l'organisme.
Le vieillissement existe pour la plupart des espèces animales et végétales, mais pas pour toutes. De très rares espèces ne manifestent aucun vieillissement observable, ou sont même capables d'inverser leur processus de vieillissement et de retourner à l'état larvaire. On peut citer notamment le krill et certains cnidaires tels que l'espèce de méduse Turritopsis nutricula et certaines hydres[1].
Le vieillissement d'un organisme débute après la phase de maturité, et progresse alors de façon irréversible jusqu'à la mort. Il se caractérise le plus souvent par une dégradation des capacités générales de l'organisme : psychomotrices, immunitaires ou reproductives.
Chez les végétaux, la sénescence peut ne toucher qu'une seule partie d'un organisme. C'est le cas de la sénescence des feuilles par exemple qui se caractérise par leur jaunissement puis leur chute en automne, ou encore des fruits lorsqu'ils tombent de la plante.
Pourquoi vieillit-on ?
La question du vieillissement a longtemps intrigué les philosophes et les naturalistes, tant il est difficile de lui trouver une explication sur le plan biologique.
Il existe un point de vue assez répandu tendant à considérer le vieillissement comme un long processus d'usure physique des tissus qui serait de même nature que l'usure de la matière inerte par le simple effet du passage du temps, c'est-à-dire par application du second principe de la thermodynamique.
D'autres approches concernent la physiologie cellulaire, notamment par l'étude des phénomènes d'oxydation (théorie mitochondriale ou théorie des radicaux libres de Denham Harman) ou d'usure des télomères (théorie télomérique du vieillissement d'Alekseï Olovnikov).
Aucune de ces explications ne supporte l'analyse détaillée, notamment compte tenu des différences de longévité qui peuvent exister entre différentes espèces.
Théories évolutionnistes

{kind=link}
Selon certaines théories évolutionnistes, le vieillissement serait une cause indirecte du fait que la longévité en elle-même n'est en général pas un critère de sélection naturelle. Un caractère procurant un avantage du point de vue de la survie ne peut pas être transmis à la descendance s'il se manifeste lorsque le cycle reproductif de l'organisme est terminé. Lorsqu'il existe en outre des facteurs pathogènes extérieurs (par exemple la prédation), un caractère favorable agissant tardivement n'a alors statistiquement que peu d'influence sur l'espérance reproductive de l'organisme. Par conséquent, les gènes agissant tard dans le développement de l'organisme ne subissent que peu de pression sélective, de telle sorte que les gènes dont l'expression est défectueuse ou délétère, sont plus nombreux lorsque cette expression intervient à un âge avancé que lorsque cette expression intervient à un jeune âge.
Cette idée est illustrée par l'observation du fait que lorsque les facteurs pathogènes extrinsèques sont absents ou inversement corrélés avec l'âge, l'espèce a d'autant plus de chances de ne pas manifester de vieillissement, comme c'est le cas pour plusieurs espèces de grandes tortues, d'esturgeons ou de grands arbres, pour lesquelles la résistance aux prédateurs augmente avec la taille et donc l'âge. Une autre illustration, pour le cas inverse, est visible chez certaines espèces dont la mort survient de façon spectaculaire après l'accouplement chez les animaux (cf. saumon), ou après la floraison chez les végétaux (cf. bambou). Ce type de reproduction est appelé « sémelparité ». La sémelparité est rarement observée parmi les mammifères, mais elle existe bel et bien, notamment chez certains marsupiaux. L'explication évolutionniste du vieillissement consiste donc dans l'idée d'une continuité entre sémelparité et itéroparité, impliquant que l'itéroparité n'est généralement pas absolue.
Une autre manière d'expliquer cette idée est de l'énoncer selon le point de vue du gène égoïste, concept imaginé par Richard Dawkins et selon lequel « une poule, ça n'est jamais que le moyen qu'a trouvé un œuf pour faire un autre œuf. » Selon ce point de vue une poule n'est pas conçue pour survivre plus longtemps qu'il ne lui est nécessaire pour pondre un œuf. Dans cette optique, l'organisme n'est qu'un outil à usage unique, un soma jetable, pour reprendre l'expression de Thomas Kirkwood, dont la seule fonction est de permettre la reproduction et la diffusion des cellules germinales.
Régulation génétique
Chez les végétaux, loin d'être une simple dégradation des conditions de vie de la cellule, voire une simple dégénérescence de celle-ci, la sénescence est un processus contrôlé génétiquement. En effet, certains gènes s'expriment uniquement au moment de la sénescence tandis que d'autres deviennent silencieux.
Sénescence cellulaire
La sénescence cellulaire a été décrite pour la première fois in vitro par Leonard Hayflick en 1961 en étudiant des fibroblastes primaires (WI-38). Il a observé qu'en culture, ces fibroblastes prolifèrent pendant un nombre limité de doublements de population avant de rester bloqués irréversiblement dans un plateau d’arrêt de croissance[2]. Ce plateau nommé limite de Hayflick ou plateau de sénescence réplicative est observé pour la plupart des cellules normales(ex. : fibroblastes, cellules endothéliales…) mais pas pour les cellules cancéreuses.
Phénotype des cellules sénescentes
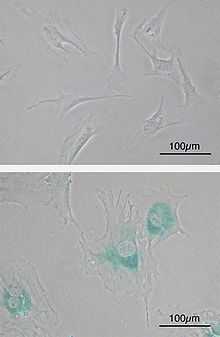
En culture, les cellules ayant atteint le plateau de sénescence présentent de nombreuses caractéristiques phénotypiques spécifiques permettant de les reconnaître aisément. Ces changements phénotypiques sont :
- augmentation de la taille des cellules et étalement sur leur support ;
- augmentation de la taille des noyaux, parfois accompagnée d’une polynucléation ;
- changements dans la structure de la chromatine (foyers d’hétérochromatine, Senescence Associated Hetero-chromatine Foci (SAHF) ;
- augmentation de l’activité lysosomiale pouvant être associé à une augmentation des activités autophagiques ;
- accumulation d’agrégats lipido-protéiques (lipofuscine) ;
- désensibilisations des cellules aux stimuli de facteurs de croissance et d’apoptose ;
- métabolisme actif permettant l’expression de transcriptomes, protéomes et sécrétomes spécifiques.
Le marquage de l’activité de la Senescence-Associated β-galactosidase (SA-βgal) est le plus couramment utilisé pour identifier des cellules sénescentes[3]. La β-galactosidase normalement active à pH 4,5 dans les lysosomes est capable de dégrader le composé X-gal dans des conditions non optimales (pH 6) dans les seules cellules sénescentes du fait de leur forte activité lysosomiale (Lee et al. 2006)[réf. insuffisante]. Bien que certaines études montrent que l’on puisse obtenir un marquage positif avec des cellules en forte confluence (Severino et al. 2000)[réf. insuffisante], le SA-βgal reste le marqueur le plus souvent utilisé pour identifier des cellules sénescentes in vitro comme in vivo.
Mécanisme moléculaire de la sénescence cellulaire
La principale conséquence moléculaire de la sénescence est un blocage du cycle cellulaire à la transition entre les phase G1 et phase S (phase de réplication de l’ADN).
Il existe 2 principales catégories de sénescence cellulaire : la sénescence réplicative et la sénescence prématurée induite par des stress. Ces stress peuvent être de natures génétiques (activation d’oncogène), métaboliques (stress oxydant) ou être environnementaux (drogues cytotoxiques). Bien qu’il y ait des similarités phénotypiques et moléculaires, les sénescences réplicative et prématurée présentent des différences importantes.
Sénescence réplicative
Sénescence induite par des stress
Sénescence et cancer
Notes et références
- ↑ (en) Absurd Creature of the Week: This Amazing Little Critter Just Might Be Immortal, Wired
- ↑ (en) Hayflick L, Moorhead PS., « The serial cultivation of human diploid cell strains », Exp Cell Res, no 25, , p. 585-621 (PMID 13905658, lire en ligne)
- ↑ (en) Dimri GP, Lee X, Basile G, Acosta M, Scott G, Roskelley C, Medrano EE, Linskens M, Rubelj I & Pereira-Smith O (1995) « A biomarker that identifies senescenthuman cells in culture and in aging skin in vivo » Proc Natl Acad Sci.U.S.A. 92:9363-7. PMID 7568133
Voir aussi
Articles connexes
- SENS Le projet initié par Aubrey de Grey qui a pour but l'extension radicale de l'espérance de vie humaine.
- Prix de la Souris Mathusalem (Methuselah Mouse Prize), une récompense pour les recherches visant à l'accroissement radical de l'espérance de vie humaine.
- Vieillesse
- Progéria, maladie présentant des signes de sénescence accélérée chez le jeune enfant
- le cas Brooke Greenberg, un défaut de vieillissement
- Cheveux blancs
Bibliographie
- Biologie de la mort, Frederic Revah et Andre Klarsfeld, Odile Jacob, ISBN 9782738107572
- Le gène égoïste, Richard Dawkins
Liens externes
|
|
Cet article ou cette section a trop de liens externes. Les liens externes doivent être des sites de référence dans le domaine du sujet. Il est souhaitable — si cela présente un intérêt — de citer ces liens comme source et de les enlever du corps de l'article ou de la section « Liens externes ».
|
(fr)
- Explications détaillées sur la sénescence cellulaire sur herman.denis.club.fr
- Chronique du livre Serons-nous immortels ? de Ray Kurzweil et Terry Grossman sur schismatrice.net
- Senescience.fr
- "Le vieillissement des cellules est réversible" sur lemonde.fr, 31 octobre 2011
- Article sur jle.com
(en)
- Mechanisms of Aging sur benbest.com
- senescence.info Informational website on aging.
- AgeLab (MIT).
- "Aging because body loses genetic info" sur meucat.com
- "The Longevity Meme (Longevity Activism)" sur longevitymeme.org
- See the artproject "Dialogue with the High Age" sur willypuchner.com
 Portail de la médecine
Portail de la médecine  Portail de la sociologie
Portail de la sociologie