
Cerebelo
Antecedentes
SOS Children han producido una selección de artículos de wikipedia para las escuelas desde el año 2005. Antes de decidir sobre el patrocinio de un niño, ¿por qué no aprender sobre diferentes obras de caridad de patrocinio primero ?
| Cerebro: Cerebelo | ||
|---|---|---|
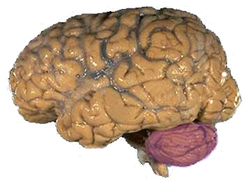 | ||
| Figura 1a: Un cerebro humano, con el cerebelo en púrpura. | ||
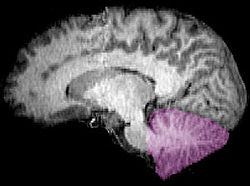 | ||
| Figura 1b: RM imagen que muestra un mediano vista sagital del cerebro humano, con el cerebelo en púrpura. | ||
| Parte de | Cerebro | |
| Arteria | SCA, AICA, PICA | |
| Vena | superior, inferior | |
El cerebelo es una región del cerebro que juega un papel importante en la integración de la percepción sensorial, la coordinación y la control del motor. Con el fin de coordinar el control motor, hay muchos caminos de los nervios que unen el cerebelo con el cerebral corteza motora (que envía información a la músculos haciendo que se mueva) y la tracto espinocerebelosa (que proporciona retroalimentación propioceptiva en la posición del cuerpo en el espacio). El cerebelo integra estas vías, como un conductor de tren, utilizando la información constante sobre la posición del cuerpo con movimientos motores afinar.
Debido a esta función 'actualización' del cerebelo, lesiones dentro de ella no son tan debilitantes como para causar parálisis, sino más bien presente como déficit de retroalimentación resultantes en los trastornos en los movimientos finos, equilibrio, postura, y aprendizaje motor. Las observaciones iniciales por fisiólogos durante el siglo 18 indican que los pacientes con problemas del cerebelo daños show con la coordinación motora y el movimiento. La investigación sobre la función cerebelosa durante la primera mitad del siglo 19 se llevó a cabo a través de estudios de lesiones y de ablación en animales . Fisiólogos investigación señalaron que tales lesiones llevaron a los animales con movimientos extraños, paso torpe, y debilidad muscular. Estas observaciones y estudios llevaron a la conclusión de que el cerebelo fue una estructura de control del motor. Sin embargo, la investigación moderna demuestra que el cerebelo tiene un papel más amplio en un número de funciones cognitivas clave, incluyendo la atención y el tratamiento de la lengua , la música y otros estímulos sensoriales temporales.
Características generales
El cerebelo se encuentra en la parte posterior inferior de la cabeza (la hindbrain), directamente a la dorsal pons, e inferior a la lóbulo occipital (Figs. 1 y 3). Debido a su gran número de pequeños células granulares, el cerebelo contiene más de 50% de todo el las neuronas en el cerebro, pero sólo ocupa el 10% del volumen total del cerebro. El cerebelo recibe casi 200 millones de fibras de entrada; En contraste, la nervio óptico está compuesto de apenas un millón de fibras.
El cerebelo se divide en dos grandes hemisferios , como el cerebro, y contiene diez lobulillos más pequeños. La citoarquitectura ( celular organización) del cerebelo es muy uniforme, con conexiones organizados en un áspero, tridimensional variedad de perpendicular elementos de circuito. Esta uniformidad de organización hace que los circuitos de los nervios relativamente fácil de estudiar. Para visualizar esta "matriz perpendicular," uno podría imaginar una calle arbolada con cables corriendo directamente a través de las ramas de un árbol a otro.
Desarrollo y evolución
{kind=link}
Durante las primeras etapas de el desarrollo embrionario, el cerebro comienza a formarse en tres segmentos distintos: la prosencéfalo, mesencéfalo, y rhombencephalon. El rombencéfalo es la más caudal (hacia la cola) segmento del cerebro embrionario; es a partir de este segmento que se desarrolla en el cerebelo. A lo largo del segmento rhombencephalic embrionario desarrollar ocho hinchazones, llamado rombómeros. El cerebelo surge a partir de dos rhombomeres ubicados en el placa alar de la tubo neural, una estructura que eventualmente forma el cerebro y la médula espinal. Los rhombomeres específicos de los que las formas cerebelo son rhombomere 1 (Rh.1) caudalmente (cerca de la cola) y el "istmo" rostral (cerca de la parte delantera).
Dos regiones primarias se cree que dará lugar a las neuronas que componen el cerebelo. La primera región es la zona ventricular en el techo de la cuarto ventrículo. Esta área produce Las células de Purkinje del cerebelo y profunda neuronas nucleares. Estas células son las neuronas de salida primarios de la corteza cerebelosa y el cerebelo. La segunda zona germinal (lugar de nacimiento celular) se conoce como el labio rómbico, las neuronas a continuación, pasar por semana embrionaria 27 a la capa granular externa. Esta capa de células que se encuentran, en el exterior del cerebelo-produce las neuronas granulares. Las neuronas granulares migran de esta capa exterior para formar una capa interna conocida como la capa de gránulos interna. La capa granular externa deja de existir en el cerebelo madura, dejando sólo las células granulares en la capa de gránulos interna. El cerebelo la materia blanca puede ser una tercera zona germinal en el cerebelo; sin embargo, su función como una zona germinal es controvertido.
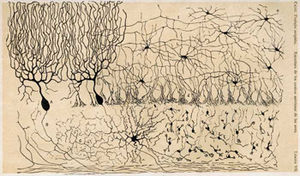
El cerebelo es de archipalliar origen filogenético. La palio es un término para la materia gris que forma la corteza. El archipallium es uno de los más evolutivamente regiones cerebrales primitivos. Los circuitos de la corteza cerebelosa aspecto similar en todos clases de vertebrados , incluyendo peces , reptiles , aves y mamíferos (por ejemplo, Fig. 2). Esto ha sido tomado como evidencia de que el cerebelo desempeña funciones importantes para todas vertebrados especies .
Anatomía
El cerebelo contiene similares divisiones de la materia gris y blanca como la cerebro. Incrustado en la materia-blanco que se conoce como el cenador vitae (Árbol de la Vida) en el cerebelo, debido a sus ramificados, treelike apariencia son cuatro núcleos profundos del cerebelo. Tres segmentos filogenéticos brutos se agrupan en gran medida por la función general. Las tres capas corticales contienen diversos tipos celulares que a menudo crean diversos retroalimentación y feedforward bucles. oxigenada sangre es suministrada por tres ramas arteriales fuera de la basilar y arterias vertebrales.
Divisiones
El cerebelo se puede dividir de acuerdo a tres criterios diferentes: anatómicas bruto, phyologenetical, y funcionales.
Divisiones anatómicas
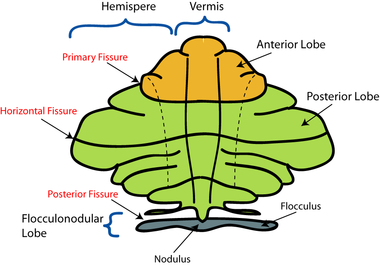
En la inspección macroscópica, tres lóbulos se pueden distinguir en el cerebelo: el lóbulo floculonodular, el lóbulo anterior (rostral a la "fisura primaria"), y el lóbulo posterior (dorsal a la "fisura primario"). Los dos últimos se pueden subdividir en una línea media vermis y hemisferios cerebelosos laterales.
 |  |
Divisiones filogenéticas y funcionales
El cerebelo también se puede dividir en tres partes sobre la base de los dos criterios filogenéticos (la edad evolutiva de cada parte) y sobre los criterios funcionales (las conexiones entrantes y salientes cada parte tiene y el papel desempeñado en la función cerebelosa normal). Desde el filogenéticamente más antigua a la más reciente, las tres partes son:
| Denominación funcional (denominación filogenético) | Piezas anatómicas | Papel |
| Vestibulocerebelo (Archicerebellum) | Lóbulo floculonodular (y vermis inmediatamente adyacentes) | El vestibulocerebelo regula los movimientos de balance y de los ojos. Se recibe información vestibular, tanto del canales semicirculares y de la núcleos vestibulares, y envía fibras de nuevo a la medial y núcleos vestibulares laterales. También recibe la información visual desde el colículo superior y de la corteza visual (este último a través de la núcleos del puente, formando una vía córtico-ponto-cerebelosa). Las lesiones de las causas vestibulocerebelo alteraciones del equilibrio y la marcha. |
| Espinocerebelo (paleocerebelo) | Vermis y partes intermedias de los hemisferios ("paravermis") | El espinocerebelo regula los movimientos del cuerpo y de las extremidades. Se recibe propiocepción de entrada de las columnas dorsales de la médula espinal (incluyendo el tracto espinocerebelar), así como de la nervio trigémino, así como de visual y sistemas auditivos. Se envía fibras a núcleos profundos del cerebelo que en el proyecto a su vez tanto a la corteza cerebral y el tronco cerebral, proporcionando así la modulación descendente de los sistemas de motor. El espinocerebelo contiene mapas sensoriales, ya que recibe datos sobre la posición de varias partes del cuerpo en el espacio: en particular, los vermis recibe fibras del tronco y las porciones proximales de las extremidades, mientras que las partes intermedias de los hemisferios reciben fibras de las porciones distales de las extremidades . El espinocerebelo es capaz de elaborar información propioceptiva con el fin de anticipar la posición futura de una parte del cuerpo durante el curso de un movimiento, de una manera "alimentar hacia adelante". |
| Cerebrocerebellum (neocerebelo, Pontocerebellum) | Partes laterales de la hemisferios | El neocerebelo está involucrado en la planificación de movimiento y la evaluación de la información sensorial para la acción. Recibe la entrada exclusivamente a partir de la corteza cerebral (especialmente el lóbulo parietal) a través de la núcleos del puente (formando vías cortico-ponto-cerebelar), y envía fibras principalmente a la ventrolateral tálamo (conectado a su vez a las áreas motoras del corteza premotora y área motora primaria de la corteza cerebral) y a la núcleo rojo (a su vez conectado a la núcleo olivar inferior, la cual remite a los hemisferios del cerebelo). El neocerebelo está involucrada en el movimiento de planificación que está a punto de ocurrir y tiene funciones puramente cognitivos. |
Gran parte de lo que se entiende acerca de las funciones del cerebelo se deriva de una cuidadosa documentación de los efectos de las lesiones focales en pacientes humanos que han sufrido una lesión o enfermedad o lesión mediante la investigación animal.
Núcleos profundos
Los núcleos profundos del cerebelo acto como los principales centros de comunicación, y los cuatro núcleos diferentes del cerebelo (dentado, interpósito, fastigial y vestibular) reciben y envían información a partes específicas del cerebro. Además, estos núcleos reciben ambas señales inhibidoras y excitadoras de otras partes del cerebro que a su vez afecta a las señales de salida del núcleo.
Capas corticales


Hay tres capas a la corteza cerebelosa; desde exterior a la capa interior, éstas son las capas moleculares, de Purkinje y granulares. La función de la corteza cerebelosa es esencialmente para modular la información que fluye a través de los núcleos profundos. El microcircuitos del cerebelo se esquematiza en la figura 5. Mossy y fibras escalada llevan información sensoriomotriz en los núcleos profundos, que a su vez la transmiten a varias áreas premotora, regulando así la ganar y el calendario de las acciones motoras. Mossy y fibras trepadoras también se alimentan de esta información en la corteza cerebelosa, que realiza diferentes cálculos, lo que resulta en la regulación de disparo de las células de Purkinje. Neuronas de Purkinje retroalimentan los núcleos profundos a través de un inhibidor potente sinapsis. Esta sinapsis regula el grado en que las fibras musgosas y escalada activan los núcleos profundos, y así controlar el efecto final del cerebelo en la función motora. La fuerza sináptica de casi todas las sinapsis en la corteza cerebelosa se ha demostrado que someterse la plasticidad sináptica. Esto permite que los circuitos de la corteza cerebelosa para ajustar continuamente y ajustar la salida del cerebelo, formando la base de algunos tipos de aprendizaje motor y la coordinación. Cada capa en la corteza cerebelosa contiene los diversos tipos de células que componen este circuito.
Capa granular
La capa más interna contiene los cuerpos celulares de dos tipos de células: los numerosos y pequeña células granulares, y el más grande Células de Golgi. Las fibras musgosas entran en la capa granular de su principal punto de origen, los núcleos del puente. Estas fibras forman sinapsis excitatorias con las células granulares y las células de los núcleos profundos del cerebelo. Las células granulares envían sus axones-conocido en forma de T como paralelo fibras-up en la capa molecular superficial, donde forman cientos de miles de sinapsis con células de Purkinje dendritas. El cerebelo humano contiene del orden de 60 hasta 80 mil millones células granulares, haciendo de este solo tipo de células, con mucho, los más numerosos de neuronas en el cerebro (aproximadamente 70% de todas las neuronas en el cerebro y la médula espinal, combinada). Células de Golgi proporcionan retroalimentación inhibidora de células granulares, formando una sinapsis con ellos y que sobresale de un axón en la capa molecular.
Capa de Purkinje
La capa intermedia contiene sólo un tipo de cuerpo que células de la gran De células de Purkinje. Las células de Purkinje son las neuronas integradoras primarias de la corteza cerebelosa y proporcionan su única salida. Dendritas de células de Purkinje son grandes pérgolas con cientos de ramas espinosas en llegar hasta la capa molecular (Fig. 6). Estas glorietas dendríticas son planas, casi todos ellos se encuentran en planos-con la vecina pérgolas de Purkinje en planos paralelos. Cada fibra paralelo desde las células granulares se ejecuta ortogonalmente a través de estos cenadores, como un alambre que pasa a través de muchas capas. Neuronas de Purkinje son GABAérgica-que significa que tienen sinapsis inhibidoras-con las neuronas del cerebelo profunda y núcleos vestibulares en el tronco cerebral. Cada célula de Purkinje recibe la entrada excitatoria de 100.000 a 200.000 fibras paralelas. Fibras paralelas se dice que son responsables por la simple (todo o nada, amplitud invariante) erizado del de células de Purkinje.
Las células de Purkinje también reciben aportaciones de la núcleo olivar inferior a través fibras de escalada. Una buena regla mnemotécnica para esta interacción es la frase "subir otro olivo", dado que las fibras trepadoras se originan de la oliva inferior contralateral. En fuerte contraste con las entradas 100,000-plus de fibras paralelas, cada célula de Purkinje recibe la entrada de fibra exactamente una escalada; pero esta sola fibra "sube" las dendritas de la célula de Purkinje, bobinado alrededor de ellos y haciendo un gran número de sinapsis que va. La entrada neta es tan fuerte que una sola potencial de acción a partir de una fibra de escalada es capaz de producir un "pico complejo" en la célula de Purkinje: un estallido de varios picos en una fila, con la disminución de la amplitud, seguido de una pausa durante la cual los picos simples se suprimen.
Capa molecular
Esta capa más externa de la corteza cerebelosa contiene dos tipos de inhibidor interneuronas: la estrellado y células en cesta. También contiene las glorietas dendríticas de las neuronas de Purkinje y tractos de fibras paralelas de las células granulares. Tanto las células estrelladas y cesta forman sinapsis GABAérgicas Onto dendritas de células de Purkinje.
Pedúnculos
Del mismo modo, el cerebelo sigue la tendencia de "tres en tres", con tres principales pedúnculos de entrada y de salida (haces de fibras). Estos son los superiores (conjunctivum brachium), media (pontis brachium), e inferior (cuerpo restiforme) pedúnculos cerebelosos.
| Pedúnculo | Descripción |
| Superior | Si bien hay algunas fibras aferentes del tracto espinocerebelar anterior que se transmitió en el lóbulo anterior del cerebelo a través de este pedúnculo, la mayoría de las fibras son eferentes. Así, el pedúnculo cerebeloso superior es la principal vía de salida del cerebelo. La mayor parte de las fibras eferentes se originan dentro de la núcleo dentado que en el proyecto a su vez a varios estructuras del cerebro medio, incluyendo el núcleo rojo, el ventral lateral / núcleo ventral anterior del tálamo, y el médula. La dentatorubrothalamocortical (dentado núcleo> núcleo rojo> tálamo> corteza premotora) y (cerebelo> tálamo> corteza premotora) itinerarios cerebellothalamocortical dos grandes vías que pasan por este pedúnculo y son importantes en la planificación motora. |
| Medio | Este se compone en su totalidad de las fibras aferentes se originan dentro de la núcleos del puente como parte de la masiva tracto corticopontocerebellar (corteza cerebral> pons> cerebelo). Estas fibras descienden de las áreas sensoriales y motoras del cerebro neocórtex y hacer que el pedúnculo cerebeloso medio la mayor de las tres pedúnculos cerebelosos. |
| Inferior | Esto lleva a muchos tipos de fibras de entrada y salida que se ocupan principalmente de la integración entrada sensorial propioceptivo con motor funciones vestibulares, tales como el equilibrio y la postura de mantenimiento. La información propioceptiva del cuerpo se lleva al cerebelo a través de la dorsal tracto espinocerebelosa. Este tramo pasa por el pedúnculo cerebeloso inferior y sinapsis dentro del paleocerebelo. Información vestibular proyecta en el archicerebellum. La fibras que suben de la carrera oliva inferior a través del pedúnculo cerebeloso inferior. Este pedúnculo también lleva información directamente del Las células de Purkinje fuera a la núcleos vestibulares en el tronco cerebral dorsal situada en la unión entre la pons y médula. |
Hay tres fuentes de entrada al cerebelo, en dos categorías consistentes en fibras musgosas y trepadoras, respectivamente. Las fibras musgosas pueden proceder de los núcleos del puente, que son grupos de neuronas localizadas en la protuberancia que llevan la información desde la corteza cerebral contralateral. Ellos también pueden surgir en el tracto espinocerebelar cuyo origen se encuentra en el ipsilateral medula espinal. La mayor parte de la salida del cerebelo sinapsis inicialmente en los núcleos cerebelares profundos antes de salir a través de los tres pedúnculos. La excepción más notable es la inhibición directa de los núcleos vestibulares de las células de Purkinje.
Suministro de sangre

Tres arterias suministran sangre al cerebelo (Fig. 7): la arteria cerebelosa superior (SCA), arteria cerebelosa anteroinferior (AICA), y arteria cerebelosa posteroinferior (PICA).
Arteria cerebelosa superior
Las ramas SCA fuera de la porción lateral de la arteria basilar, justo por debajo de su bifurcación en la arteria cerebral posterior. Aquí se envuelve alrededor de la protuberancia posterior (a la que también suministra sangre) antes de llegar al cerebelo. El SCA suministra sangre a la mayor parte de la corteza cerebelosa, los núcleos del cerebelo, y los pedúnculos cerebelosos medios y superiores.
Arteria cerebelosa inferior anterior
Las ramas AICA fuera de la porción lateral de la arteria basilar, justo por encima de la unión de las arterias vertebrales. Desde su origen, se ramifica a lo largo de la parte inferior de la protuberancia en el ángulo pontocerebeloso antes de llegar al cerebelo. Esta arteria suministra sangre a la porción anterior del cerebelo inferior, y para la facial (NC VII) y nervios vestibulococlear (CN VIII).
La obstrucción de la AICA puede causar paresia, parálisis y pérdida de sensibilidad en la cara; también puede causar daños auditivos . Además, podría causar un infarto del ángulo pontocerebeloso. Esto podría llevar a hiperacusia (disfunción del músculo estapedial, inervado por CN VII) y vértigo (interpretación equivocada de la vestibular del canal semicircular de endolinfa aceleración causada por la alteración de CN VIII).
Arteria cerebelosa posteroinferior
Las ramas PICA fuera de la parte lateral de las arterias vertebrales justo por debajo de su unión con la arteria basilar. Antes de llegar a la superficie inferior del cerebelo, la PICA envía ramas en la médula, el suministro de sangre a varios núcleos de los nervios craneales. En el cerebelo, la PICA suministra sangre a la parte posterior inferior del cerebelo, el pedúnculo cerebeloso inferior, la núcleo ambiguo, el núcleo motor vago, la médula núcleo del trigémino, la núcleo solitario, y la núcleos VESTIBULOCOCLEAR.
Función general
Funcionalmente, el fibra de la escalada y el vías de fibra de células paralelas de fibra de gránulos cubiertos de musgo son los dos tipos principales de los aferentes al cerebelo en su conjunto y de las células de Purkinje, en particular. Estos sistemas aferentes difieren drásticamente en su conectividad. La célula de Purkinje y su fibra aferente escalada tienen una relación uno-a-uno y la proyección global está organizada para producir la activación sincrónica de grupos específicos de células de Purkinje en una orientación rostrocaudal. La relación entre la célula de Purkinje y el sistema de fibras de fibra paralela musgo puede caracterizarse como de muchos a muchos. Con la direccionalidad siendo orientación mediolateral dentro de la capa molecular, es decir, en ángulo recto con las dendritas de células de Purkinje que son isoplanar.
El sistema de fibra escalada
Se origina a partir de la oliva inferior contralateral. Como resultado del acoplamiento eléctrico entre las neuronas olivar inferior, su desacoplamiento dinámico través de la inhibición de retorno de los núcleos cerebelares y la topografía de la proyección olivocerebellar, este sistema genera síncrona (en una escala de tiempo de milisegundos) la activación pico compleja de células de Purkinje, en rostrocaudally bandas orientadas. Estas bandas de actividad son alrededor de 250 micras de ancho en la dirección mediolateral pero pueden ser de varios milímetros de longitud en la dirección rostrocaudal y extenderse hacia abajo las paredes de la folia cerebelosa y a través de varios lóbulos. La distribución de la sincronía de momento a momento del control motor se modula de forma dinámica por la oliva inferior con el importante papel de los aferentes olivares son para determinar el patrón de acoplamiento electrónico "efectiva" entre las neuronas olivares y con ello la distribución de la actividad pico complejo sincrónico, entre la corteza cerebelosa. Cambios en los patrones de sincronía están asociados con los traslados de animales que realizan una tarea de motor .. En efecto. El sistema olivocerebellar puede considerarse un sustrato eléctricamente maleable desde el que las sinergias de motor único se puede esculpir.
El sistema de fibra Mossy Fiber-Parallel
En contraste con la naturaleza punteada de la activación cerebelosa por el sistema olivocerebellar, el sistema de fibras de fibra paralelo de musgo proporciona una regulación continua y muy delicado de la excitabilidad de los núcleos cerebelares , provocada por la activación tónica de picos simples en las células de Purkinje, que en última instancia genera el control fino del movimiento conocido como la coordinación motora. El hecho de que las fibras musgosas informan la corteza cerebelosa de tanto ascendente como descendente mensajes hacia y desde los centros motores en la médula espinal y el tronco cerebral nos da una idea del papel fundamental del sistema de fibras musgosas: se informa a la corteza del lugar y velocidad de movimiento de las extremidades y pone las intenciones de motor generados por el cerebro en el contexto de la situación del cuerpo en el momento del movimiento se va a ejecutar. Por otra parte, a través de sus efectos sobre las células inhibidoras GABAérgicas cerebelo nucleares, que se proyectan de nuevo a la oliva, ayuda a dar forma a la estructura de acoplamiento entre las células olivares y por lo tanto la distribución de sincronía en la próxima aprobación de la gestión olivocerebellar.
El cerebelo Núcleos
Las células de Purkinje son la única salida de la corteza cerebelosa y son de naturaleza inhibidora Sus axones en contacto con el cerebelosa y Deiters núcleo vestibular como su único objetivo. La actividad de los núcleos cerebelares se regula en tres maneras: (1) por la entrada excitadora de colaterales de los sistemas aferentes cerebelosas, (2) por las entradas inhibidoras de las células de Purkinje activados sobre las vías de fibras musgosas, y (3) por las entradas de Purkinje células activadas por el sistema de fibras de escalada
Función general cerebelosa
La salida del cerebelo (los axones de los núcleos cerebelares) proceder para generar la actividad de fondo que sirve para ajustar el tono general y la postura que le da la corteza motora la capacidad de ejecutar movimientos sobre la base de la intención (la estrategia de movimiento). En este contexto, el cerebelo ofrece la táctica de la activación muscular múltiple requerida para soportar tales definen movimientos. Y así, mientras que el cerebro del motor determina dónde se mueven (imperativo ejecutivo) el cerebelo implementa su momento adecuado y modula la fuerza dada a todos los comandos del motor, como la coordinación de movimiento es una función no continua.
Disfunción
Ataxia es un complejo de síntomas, por lo general implica una falta de coordinación, que se encuentra a menudo en los procesos de enfermedad que afectan el cerebelo. Para identificar los problemas del cerebelo, los examen neurológico incluye una evaluación de la marcha (una marcha de base amplia que es indicativa de la ataxia), pruebas en forma de dedo que señala y evaluación de la postura. Las anomalías estructurales del cerebelo (hemorragia, infarto, neoplasia, degeneración) se pueden identificar en las imágenes de la sección transversal. La resonancia magnética es la técnica de elección, como La tomografía computarizada es suficientemente sensible para detectar anomalías estructurales del cerebelo.
Envejecimiento
La estudio estereológico ha encontrado que cerebelosa humana la materia blanca se redujo en un 26% con edad (en el rango de edad 19 a 84). Los investigadores del estudio no pudo detectar pérdida global de Purkinje o células granulares, sin embargo en el lóbulo anterior hubo una pérdida significativa de estos tipos de células, así como una pérdida de volumen del 30%. Con imágenes por resonancia magnética una reducción volumétrica moderada con la edad en vermis y hemisferio cerebeloso se ha observado.
Una estudio autorradiografía del cerebelo humano encontró una creciente unión de H-3- ketanserin con la edad. (Ketanserina se une principalmente a la 5-HT 2A neurorreceptor) El mismo equipo de investigación no encontró ninguna correlación significativa con la edad en su homogenizar estudio vinculante. Un tanto de acuerdo con el estudio autorradiografía un estudio de tomografía por emisión de positrones con la altanserin receptor 5-HT 2A radioligando encontró una correlación positiva entre la edad y cerebelosa unión no específica.
Las teorías sobre la función cerebelosa
Dos teorías Dirección principal de la función del cerebelo, tanto frente a la coordinación motora. Uno afirma que las funciones del cerebelo como un regulador del "tiempo de los movimientos". Esto se desprende de los estudios de pacientes cuyos movimientos cronometrada se ven perturbadas.
El segundo, "Teoría de Redes Tensor" ofrece un modelo matemático de transformación de los sentidos (covariante) coordenadas espacio-temporales en el motor (contravariante) coordina las redes neuronales del cerebelo.
Al igual que muchas controversias en las ciencias físicas, pero existe evidencia de cada una de las hipótesis anteriores. Los estudios sobre el aprendizaje motor en el reflejo vestíbulo-ocular y Condicionamiento del parpadeo demostrar que el momento y amplitud de movimientos aprendidos están codificadas por el cerebelo. Muchos mecanismos de plasticidad sináptica se han encontrado en todo el cerebelo. El modelo de Marr-Albus atribuye principalmente aprendizaje motor a un solo mecanismo de plasticidad: el depresión a largo plazo de las sinapsis de fibras paralelas. La Teoría Tensor Red de transformaciones sensoriomotores por el cerebelo también se ha apoyado de forma experimental.
Con la llegada de más sofisticado técnicas de neuroimagen tales como La tomografía por emisión de positrones (PET), y fMRI, numerosas y diversas funciones están al menos parcialmente atribuidos al cerebelo. Lo que antes era piensa que es principalmente un / región de integración sensorial motor está demostrando estar involucrado en muchas diversas funciones cognitivas.
Modelado cerebelosa
Como se ha mencionado en la sección anterior, ha habido muchos intentos de modelar la función del cerebelo. Los conocimientos proporcionados por los modelos también han dado lugar a extrapolaciones en los dominios de metodologías de inteligencia artificial, especialmente redes neuronales. Algunos de los logros más notables han sido Cerebellatron, cerebelosa Modelo redes CMAC memoria asociativa o, y SpikeFORCE para control de movimiento robótico, y "Teoría de Redes Tensor".
Imágenes adicionales
- TC del cerebro de Mikael Häggström S3 I8.JPG
La tomografía computarizada de la cabeza, con el cerebelo visible en la parte inferior.

Lóbulos

Diencephalon
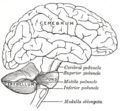
Esquema que muestra las conexiones de las diversas partes del cerebro.

La superficie superior del cerebelo.

Bajo superficie del cerebelo.

Corte sagital del cerebelo, cerca de la unión del vermis con el hemisferio.

La disección que muestra las fibras de proyección del cerebelo.

Esquema del techo del cuarto ventrículo. La flecha está en el agujero de Majendie.

La disección que muestra el curso de las fibras cefalorraquídeo.

Diagrama que muestra las posiciones de los tres cisternas subaracnoidea director.
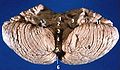
Cerebelo humano vista anterior
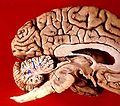
Cerebro humano vista sagital medio